一个涉及PPARγ、酰基辅酶A结合蛋白和GABAA受体的肥胖促进前馈环
酰基辅酶A结合蛋白(ACBP),也称为地西泮结合抑制剂(DBI),是食欲和脂肪生成的强效刺激物。生物信息学分析结合系统筛选揭示,过氧化物酶体增殖物激活受体γ(PPARγ)是最佳解释ACBP/DBI在包括肝脏和脂肪组织在内的代谢活跃器官中上调的转录因子。PPARγ激动剂罗格列酮诱导ACBP/DBI上调,以及体重增加,这可以通过在小鼠中敲除_Acbp_/Dbi_来预防。此外,肝脏特异性敲低_Pparg_防止了高脂饮食(HFD)诱导的循环ACBP/DBI水平上调,并减少了体重增加。相反,敲除_Acbp/_Dbi_防止了HFD诱导的PPARγ上调。值得注意的是,γ-氨基丁酸A受体(GABAAR)γ2亚基中的单个氨基酸替换(F77I),使ACBP/DBI与该受体的结合消失,防止了HFD诱导的体重增加,以及HFD诱导的ACBP/DBI、GABAAR γ2和PPARγ的上调。基于这些结果,我们推测存在一个依赖于ACBP/DBI、GABAAR和PPARγ的肥胖前馈循环。在任何层面中断这个恶性循环,都会明显减轻HFD诱导的体重增加、肝脂肪变性和高血糖。
文章 开放获取 2024年11月15日
文章 开放获取 2021年1月14日
肥胖已成为全球最普遍的病理状态,其合并症(包括糖尿病、心血管疾病和癌症)占据了整体死亡率中日益增加的份额 [1, 2]。尽管肥胖的病因复杂,但它似乎构成了一种接近不可逆转的状态,使越来越不健康的患者陷入了过度饮食的永久习惯。尽管有饮食、运动、生活方式干预、药物甚至外科手术,但绝大多数患者仅表现出短暂的体重减轻,随后出现反弹效应。因此,国家体重控制登记处(包括至少减重13.6公斤并持续至少1年的成年人,http://www.nwcr.ws)在美国大约1亿肥胖人口中仅有10,000名成员(即每10,000名肥胖个体中仅有1名)。
多种交织的遗传、心理社会、神经精神、神经内分泌、代谢、炎症、免疫,甚至微生物组基础的回路被认为与肥胖相关的食物成瘾有关 [3,4,5]。显然,许多研究旨在识别肥胖个体中食欲刺激因子的增加——或食欲抑制因子的缺乏。然而,令人惊讶的是,大多数已知的食欲刺激因子(如胃饥饿素)在肥胖者中实际上是减少的 [6],而大多数食欲抑制因子(如瘦素)在非综合征性肥胖中是增加的 [7],这可能反映了一种失调的稳态调节状态 [8]。只有少数食欲刺激因子在肥胖个体中真正升高,例如由基因 diazepam-binding inhibitor (DBI) 编码的酰基辅酶A结合蛋白(ACBP) [9,10,11]。
ACBP/DBI(以下简称 ACBP)是一种系统发育上古老的蛋白质,通常存在于有核细胞的细胞质中,在那里它与活化的中链脂肪酸相互作用并参与脂质代谢 [12, 13]。作为一种无领导肽,它不经历传统的蛋白质分泌,然而,它可以通过与自噬相关的途径释放到细胞外空间 [9, 14]。因此,禁食1到2天的老鼠表现出循环 ACBP 蛋白水平的自噬依赖性增加 [9],而经历自愿禁食数周(在减肥诊所)或非自愿禁食(由于癌症化疗)的人类受试者也显示出血浆 ACBP 的升高 [11]。在饥饿的老鼠中,通过适当的抗体中和 ACBP 会显著减少再喂养,表明 ACBP 作为一种真正的食欲刺激物 [9]。实际上,静脉注射 ACBP 会引发进食行为,其在肝脏中的转基因过表达会导致体重增加 [10]。类似地,在线虫 Caenorhabditis elegans 和果蝇 Drosophila melanogaster 中,哺乳动物 ACBP 的同源物分别刺激咽部泵动和口钩运动,这与 ACBP 是一种系统发育上古老的食物摄入刺激物的观点一致 [15, 16]。总的来说,现有证据表明,在老鼠中,ACBP 嵌入在一个电路中,饥饿刺激自噬,从而导致 ACBP 从细胞释放。分泌的 ACBP 作用于表达在中枢神经系统外的多种细胞类型(例如,胆管细胞、肝细胞和巨噬细胞)上的 γ-氨基丁酸 A 受体(GABAAR) [17,18,19],导致血糖的短暂下降和随之而来的食欲刺激电路的激活 [9,10,11],从而刺激食物摄入并关闭一个体内稳态反馈回路或“饥饿反射” [20]。
循环中的 ACBP 水平在肥胖小鼠和人类中升高。在人类中,血浆 ACBP 与体重指数 (BMI) 相关,并且与升高的心脏代谢风险因素相关,如血糖、空腹胰岛素水平、总胆固醇、甘油三酯、肝转氨酶和收缩压 [9,10,11],以及全身炎症的迹象 [21]。血浆 ACBP 的增加与肥胖相关的肝脏和白色脂肪组织 (WAT) 中 ACBP mRNA 的上调(在小鼠和患者中)以及肥胖小鼠的肝脏和 WAT 中 ACBP 蛋白的上调相关 [9]。相反,人类厌食症伴随着 ACBP 血浆水平的降低 [9, 22]。因此,长期的 BMI 变化似乎与“饥饿反射”的稳态丧失相关,导致系统设定点的永久性和病理性改变。在一个假设的肥胖前馈循环中,过度饮食会导致 ACBP 水平的增加,而这又促进过量的食物摄入 [23]。
受到上述假设的启发,我们决定识别那些最佳解释与肥胖相关的基因表达谱变化的转录因子(TFs),包括ACBP的上调。在这里,我们报告过氧化物酶体增殖物激活受体伽马(PPARγ)在响应肥胖刺激(包括高脂饮食(HFD)和PPARγ激动剂)时,作用于编码ACBP的基因进行转激活。有趣的是,ACBP还通过其对GABAAR的作用刺激PPARγ的活性,从而完成一个可能有助于维持肥胖状态的恶性放大循环。
培养基和细胞培养补充剂购自Gibco-Invitrogen(美国加利福尼亚州卡尔斯巴德)。塑料器皿购自Corning B.V. Life Sciences(荷兰斯基普霍尔-瑞克)。除非另有报告,本研究中使用的所有细胞系均在标准条件下培养(37 °C,5% CO2,含10%胎牛血清的Dulbecco改良的Eagle培养基,10 mM HEPES缓冲液,100 U/mL青霉素G钠和100 mg/mL链霉素硫酸盐)。人肝细胞癌(HepG2,ATCC #HB-8065)、小鼠肝细胞癌(Hep55.1c,ATCC)和小鼠肝癌(Hepa1–6,ATCC #CRL-1830)细胞系用于体外实验。细胞系经过检测未发现支原体污染。细胞被接种在6孔、12孔或9孔板中,并在处理前培养24小时。HepG2细胞每日用PPARγ激动剂罗格列酮(2.5 μM,Sigma,美国密苏里州圣路易斯,#R2408)、依达列酮(2.5 μM,Tocris Bio-Techne,英国布里斯托,#4784)、GW1929(2.5 μM,Sigma,美国密苏里州圣路易斯,#G5668)、S26948(2.5 μM,Sigma,美国密苏里州圣路易斯,#SML0510)处理,并在8天后收获以通过细胞荧光计测定评估细胞内ACBP蛋白水平。此外,HepG2细胞用PPARγ激动剂罗格列酮(2.5 μM)和依达列酮(5.0 μM)处理7天,然后进行针对_PPARG_的_siRNA_转染。转染后96小时,通过细胞计数法定量ACBP水平。
稳定 shAcbp 表达的 Hep55.1c 细胞系
针对 Acbp 的 shRNA (shAcbp-1, TRCN0000317379; shAcbp-2, TRCN0000105052; shAcbp-3, TRCN0000105050),以及阴性对照 shRNA 被克隆到 pLKO.1-puro 逆转录病毒载体中(Sigma, Burlington, MA, USA)。约 2 × 105 Hep55.1c 细胞被接种在六孔板中(DMEM, 10% FBS),直到 60–70% 融合度。接下来,细胞用 lenti-shRNA 粒子(25–35 μL)在 1 mL 新鲜培养基(DMEM, 10% FBS, 5 μg/mL Polybrene, Sigma, Burlington, MA, USA, # TR-1003)中感染。转染后 24 到 48 小时,培养基被更换(DMEM, 10% FBS),细胞再培养一天。感染的 Hep55.1c 细胞在 1 周内被选择(10 µg/mL,嘌呤霉素,Thermo Fisher Scientific, Carlsbad, CA, USA, #A1113803)。为了单克隆稳定细胞系的分离,按照标准协议进行了单细胞分选。通过 Acbp 定量 PCR 和免疫印迹法确定 lenti-shRNA 对 Acbp 的效率。为了功能分析,阴性对照和 shRNA-Acbp Hep55.1c 单克隆细胞系被用罗格列酮处理 48 小时,随后进行 ACBP 和 PPARγ 的免疫印迹。
细胞荧光测定
细胞使用 Accutase (StemPro Accutase 细胞解离试剂, Thermo Fisher Scientific, Carlsbad, CA, USA, #A1110501) 收集,并在用 2% PFA 的 PBS 固定 20 分钟后用 PBS 洗涤两次。细胞用 0.1% Triton X-100 渗透 10 分钟,随后用冷封闭溶液 (3% BSA, v/v 在 PBS 中) 洗涤两次,并在 4 °C 下与一抗孵育过夜。细胞被洗涤并在封闭缓冲液中与二抗 AlexaFluor 647-结合物孵育 (60 分钟),然后在流式细胞仪分析前洗涤,使用 MACSQuant 细胞计数仪 (Miltenyi Biotec, Bergisch Gladbach, Germany)。
沉默 RNA (siRNA) 在体外的敲低
'siRNAs 使用 Lipofectamine™ RNAiMAX 转染试剂 (Invitrogen, Carlsbad, CA, USA, #13778150) 根据制造商的说明进行转染,使用 100 nM 的针对 ACBP、ACOT1(四个序列:ACOT1、ACOT2、ACOT3、ACOT4)、PPARG(五个序列:PPARG Ambion、PPARG i、PPARG ii)以及针对 CTCF、POLR2A、E2F1、SP1 NANOG、NFYA、EP300、FOS、NR3C1、CEBPB、MYC、FOXA1、FOXA2、TAF1、PHF8、JUND、MTA3、FOXP2、MBD4、PPARG 和 SREBP 的 SMARTpool siRNAs 进行 96 小时的孵育。细胞按照描述进行处理,并进行下游分析。使用空载体或无关的 siRNA (UNR) 作为对照.
免疫印迹
约 20–25 μg 的蛋白质裂解液通过 SDS-PAGE 在 4–12% Bis-Tris 丙烯酰胺预制凝胶 (Thermo Fisher Scientific, Carlsbad, CA, USA, #WG1402BOX) 中分离,并电转移到免疫印迹 PVDF 膜 (Biorad, Hercules, CA, USA, #1620177)。为了评估电转移效率,膜用 Ponceau S 溶液染色,然后在 TBST 溶液中冲洗和清洗。将膜在封闭缓冲液中孵育 2 小时,以饱和非特异性结合位点。封闭缓冲液在 TBST 溶液中冲洗,膜在一抗中孵育过夜 (人 ACBP: Santa-Cruz #sc-376853,鼠 ACBP: Abcam #ab231910,鼠 FASN: Cell Signaling Technology #3180,鼠 GABRG2: Abcam # ab87328,鼠 GAPDH: Cell Signaling Technology #2118,鼠 PPARγ: Abcam #ab178860 & Santa-Cruz #sc-7273)。然后在 TBST 中清洗膜,接着在辣根过氧化物酶 (HRP) 标记的二抗混合物 (Southern Biotech, Birmingham, USA) 中孵育。最后在 TBST 溶液中清洗膜,并使用 SuperSignal West Pico 化学发光底物 (Thermo Fisher Scientific, Carlsbad, CA, USA, #34579) 通过 ImageQuant™ LAS 4000 (GE Healthcare) 显影。
共同免疫沉淀实验
ACBP与GABAAR γ2亚基之间的物理相互作用通过标准免疫沉淀(IP)和免疫印迹协议进行了研究。具体而言,肝脏蛋白提取物(500 μg)在蛋白A/G-Sepharose珠子(Merck Millipore, Burlington, MA, USA, #GE17–0618–01)上使用山羊多克隆GABAAR γ2抗体(每个IP反应3 μg,Abcam #ab240445)或其阴性同型对照(IgG)进行免疫沉淀。每个IP反应在旋转室中孵育过夜(4 °C),随后在第二天进行了三轮连续的PBS洗涤。每轮洗涤包括对沉淀的PBS重悬和重新离心(12,000×g, 4 °C)。最后,珠子在20 μL的NUPAGE 4x缓冲液(Life Technologies, CA, USA, #NP0008)中重悬,100 °C加热(10 分钟),然后进行标准免疫印迹以检测GABAAR γ2蛋白和感兴趣的蛋白。
小鼠实验
本研究中使用的所有小鼠均在无病原体、温控环境中饲养和居住,光暗周期为12小时,符合FELASA指南、欧盟指令63/2010和法国立法。Acbp__fl/fl、Pparg__fl/fl_和_Gabrg2tm1Wul/J 8-12周龄雄性小鼠在内部繁殖(CEF,法国巴黎)。C57BL/6 8-12周龄雄性小鼠购自Envigo(Envigo,法国Gannat)。B6.V-Lep__ob/ob__JRj(肥胖)和_B6.V-Lep__ob/T__JRj_(瘦)8-12周龄雄性小鼠购自Charles River(Charles River Laboratory,法国Lentilly)。所有小鼠均自由获取常规(Safe,#A04)或高脂肪饲料(Safe,#260 HF)和水。在药理学研究中,罗格列酮、贝克拉酮(Sigma,圣路易斯,MO,美国,#SML0282)和HX531(Sigma,圣路易斯,MO,美国,#SML2170)分别以6、25和33 mg/kg体重的剂量每日腹腔注射给_C57BL/6_小鼠,持续5天。除非另有报告,体重每周监测一次。所有动物实验均遵循“C. Darwin”地方动物实验伦理委员会的规定(协议#25355,# 10862,# 8530,#25010,# 0447.02,#25000)。
脂肪细胞特异性 Acbp 敲除小鼠
脂肪组织特异性 Acbp 基因敲除小鼠的生成如前所述 [11]。简而言之,Acbp__fl/fl 小鼠(美国华盛顿州本特利)与 B6;FVB-Tg(AdipoQ-cre)1Evdr/J 小鼠(杰克逊实验室)交配。
条件性全身 Pparg 敲除小鼠
全身PPARγ敲除小鼠是通过将_Pparg__fl/fl_小鼠(Pparg__tm2Rev,loxP位点夹在外显子1和2之间;Bentley, WA, USA)与_B6.Cg-Tg_ (UBC-cre/ERT2)1Ejb/1 _J_小鼠(Jackson Laboratory, Bar Harbor, ME, USA)交配生成的。在繁殖过程中,通过使用从尾部活检中分离的基因组DNA进行PCR,使用针对_Cre_和_Pparg_基因位点的特异性引物验证基因型。
Pparg 引物:
- (1)
_PpargF_ (_oIMR1935_), 5′ TGGCTTCCAGTGCATAAGTT 3′
- (2)
_PpargR_ (_oIMR1934_), 5′ TGTAATGGAAGGGCAAAAGG 3′
PCR 条件:95 °C (5 min); [95 °C (30 s), 60 °C (30 s), 72 °C (45 s)] x 35 个循环; 72 °C (5 min)。PCR 产物通过 2% 琼脂糖凝胶电泳分离,使用溴化乙锭溶液 (Sigma, #E1510) 可视化,或使用毛细管电泳系统 LabChip® GX Touch™ (PerkinElmer)。
Cre 引物:
- (1)
_Cre1_, AGGTTCGTTCACTCATGGA;
- (2)
_Cre2_, TCGACCAGTTTAGTTACCC.
PCR 条件:95 °C (5 min); [95 °C (30 s), 60 °C (30 s), 72 °C (45 s)] x 35 个循环; 72 °C (5 min)。PCR 产物通过 2% 琼脂糖凝胶电泳分离,使用溴化乙锭溶液 (Sigma, #E1510) 可视化,或使用毛细管电泳系统 LabChip® GX Touch™ (PerkinElmer)。
Cre重组酶的表达在给予他莫昔芬后被诱导(以75 mg/kg的体重通过腹腔注射的方式每日一次,持续5天)。他莫昔芬之前在玉米油(90%)和乙醇(10%)中稀释至最终浓度为20 mg/ml,然后在37 °C下孵育过夜。
GABAAR γ2 亚基突变小鼠
'Gabrg2tm1Wul/J 本研究使用了 8–12 周龄的雄性小鼠 [24]。这种遗传背景编码了 γ-氨基丁酸 A 受体 (GABAAR) γ2 亚单位中的一个点突变 (F77I) (JAX™ 小鼠品系,查尔斯河实验室,法国伦蒂利),在纯合状态下,导致 ACBP-GABAAR 结合在全身范围内受到影响。'
在繁殖期间,通过使用从尾部活检中分离的基因组DNA和特异于_Gabrg2_基因位点的引物进行PCR验证基因型。
Gabrg2 引物:
- (1)
_Gabrg2F (16697)_, 5′ AAGCGCCCACCTCTACTTCT 3′
- (2)
_Gabrg2R (16698)_, 5′ TCATGGGATAGTGCATCAGC 3′
PCR 条件:95 °C (5 min); [95 °C (30 s), 60 °C (30 s), 72 °C (45 s)] x 35 个循环; 72 °C (5 min)。PCR 产物通过 2% 琼脂糖凝胶电泳分离,使用溴化乙锭溶液 (Sigma, #E1510) 可视化,或使用毛细管电泳系统 LabChip® GX Touch™ (PerkinElmer)。
ACBP 中和在体内的作用
小鼠被腹腔注射小鼠同种型IgG(阴性对照;2.5 μg/g 体重,200 μL)或单克隆抗ACBP中和抗体(2.5 μg/g 体重,200 μL),每周两次,持续6周(inVivoMAb #C1.18.4_BE0085 和 Fred Hutch Antibody Technology #N/A)。
单导RNA (sgRNA) 序列的设计与分子克隆
sgRNA寡核苷酸序列(针对 Pparg 外显子 2: 5′ GTTCATGAGGCCTGTTGTAGAG 3′,以及阴性对照 Rosa26 内含子 1: 5′ GCTCGATGGAAAATACTCCGAG 3′)被设计(Broad Institute, portals.broadinstitute.org/gpp/public/analysis-tools/sgrna-design),合成(Eurogentec)并克隆到 pX602-TBG 质粒载体中(BsaI 限制酶)(Addgene, #61593) [25]。成功克隆通过 DNA Sanger 测序(Eurofins, U6 引物)验证。
肝脏特异性 CRISPR/Cas9 PPARγ 基因敲除
'Rosa26 内含子 1 基因位点突变作为阴性对照 (Loesch et al. bioRxiv, 2021)。AAV8 载体 (rAAV 平台,Imagine institute,法国) 通过眼眶后注射给 4 周龄雄性 C57/Bl6 小鼠。所有 AAV8 剂量调整为 150 µL,使用无菌生理盐水稀释至每只小鼠 2 × 1011 vg 的浓度。小鼠被随机分配到不同的实验条件。突变在肝细胞分离物中通过 DNA 水平验证 (Pparg 外显子 2 或 Rosa26 位点 PCR 扩增,Sanger 测序,Eurofins),并使用 TIDE 在线软件分析 indel 百分比 (https://tide.nki.nl/).'
'Pparg 外显子 2 引物:'
- (1)
_PpargF_, 5′ TTGTGCAGGTGAGGTTCTGG 3′
- (2)
_PpargR_, 5′ ACAGACTCGGCACTCAATGG 3′
'Rosa26 内含子 1 引物:'
- (1)
_Rosa26F_, 5′ CTTGCTCTCCCAAAGTCGCT 3′
- (2)
_Rosa26R_, 5′ CCAATGCTCTGTCTAGGGGT 3′
PCR 条件:95 °C (5 min); [95 °C (30 s), 55 °C (30 s), 70 °C (30 s)] x 30 个循环; 70 °C (10 min); 4 °C。PCR 产物通过 2% 琼脂糖凝胶电泳分离,并使用 SYBR Safe DNA 凝胶染料(Invitrogen, Carlsbad, CA, USA, #S33102)进行可视化。
'Pparg 突变在蛋白质水平上通过免疫印迹进一步验证(Abcam,剑桥,英国,#ab178860)。'
肝脏组织学
小鼠肝脏样本在20 mL 4% v/v 甲醛溶液(4 °C)中固定24小时,随后进行脱水(在逐渐增加的乙醇溶液中孵育;70–100% v/v)和石蜡包埋。五微米切片使用苏木精-伊红(HE)染色,并通过蔡司Lame Axioscan扫描(物镜:×20)。图像使用Zen软件进行分析。
NAFLD 评分测量
NAFLD 活动评分是根据标准协议进行测量的。简而言之,计算描述脂肪变性(0–3)、肝细胞气球样变(0–2)和小叶炎症(0–3)的数值评分之和 [26]。
ACBP 和小鼠血浆样本中的瘦素检测
小鼠血液通过亚颌静脉使用锂肝素血液采集管(Sarstedt, France, #16443)获得,随后立即进行离心(12,000×g, 30 min, 4 oC)以分离血浆。血浆在冰冷的PBS中稀释1:20,并根据制造商的说明用于ACBP(MyBioSource, #MBS2025156)和瘦素(Merck Millipore, Burlington, MA, USA, #EZML-82K)定量。
食物摄入测量
八周大的雄性小鼠被单独饲养,并在25 × 15 × 10 cm的笼子中适应,随后在实验前进行了6–8小时的禁食(期间自由饮水)。在12小时的过程中监测了累计食物摄入量。
血糖(血糖值)和β-羟基丁酸测量
空腹(12小时)血糖使用Accu-Chek Performa机器(Accu-Check,法国,#4702354)测量。空腹(12小时)血β-羟基丁酸水平使用Glucofix Ketone B读数器(Menarini,佛罗伦萨,意大利,#MEN45800)测量。
代谢组学分析(组织和血浆)样本准备
从每只牺牲的小鼠中收集约30 mg的肝脏,放入一个2-mL的匀浆管(硬组织匀浆CK28,2.8 mm氧化锆珠;Precellys,Bertin Technologies,法国),其中含有1 mL冰冷的提取混合物(甲醇/水,9/1,−20 °C,含有内标的混合物)。样品经过匀浆(三个循环,每个循环20 秒/ 5000 rpm;Precellys 24,Bertin Technologies,Montigny-le-Bretonneux,法国),以促进溶剂的接触和内源性代谢物的提取,匀浆液经过离心(在15,000×g,4 °C下离心10 分钟),最后收集上清液。或者,将血浆样品(25 μL)与250 µL冰冷的提取混合物混合,允许蛋白质沉淀和代谢物提取,震荡后离心(在15,000×g,4 °C下离心10 分钟)。
在样品离心后(无论是来自器官组织还是血浆),收集上清液,分成三个部分,并按照标准协议处理 [27]。简而言之,第一个部分用于短链脂肪酸的下游分析(组织和血浆样品各40 µL),第二个部分用于液相色谱/质谱(LC/MS)工作流程,第三个部分用于气相色谱/质谱(GC/MS)工作流程分析。每个组织约300 µL,每个血浆样品约100 µL被转移到注射琥珀色玻璃瓶(带有熔融插入物)中,并在40 °C下蒸发(Techne DB3,英国斯塔福德郡)。第二个干燥部分用200或150 µL(分别为组织或血浆样品)的超纯水恢复,并在−80 °C下储存,直到通过LC/MS注射和分析。第三个干燥部分在GC/MS注射和分析之前进行了衍生化。最后,第四个部分和样品沉淀用等体积的2% SSA(在MeOH中)重新提取,涡旋混合并离心(在15,000×g,4 °C下10分钟)。上清液(分别来自组织和血浆提取物的350和60 µL)被转移到注射聚丙烯瓶(带有熔融插入物)中,并在40 °C下蒸发(Techne DB3,英国斯塔福德郡)。干燥样品用超纯水恢复(分别为组织和血浆,干燥提取物200和100 µL),并在−80 °C下储存,直到通过(超高效液相色谱/质谱)UHPLC/MS进行多胺检测。
超高效液相色谱/质谱
针对性的 UHPLC/MS 分析使用 UHPLC 1290 系统(安捷伦科技,德国瓦尔德布龙)进行,包括一个自动进样器(4 °C)和一个佩尔蒂尔烘箱,以严格控制柱温。在 UHPLC 上耦合了三重四极杆质谱仪(QQQ/MS)6470(安捷伦科技),配备电喷雾源,使用氮气作为碰撞气体。通过将 10 μL 样品注入 Zorbax Eclipse XDB-C18(100 mm × 2.1 mm,粒径 1.8 µm;安捷伦)柱,检测到短链脂肪酸和酮体,保护柱为 C18(5 mm × 2.1 mm,粒径 1.8 μm)。分析期间,柱烘箱保持在 50 °C。梯度流动相由 0.01% 甲酸(西格玛)(A)和 ACN(0.01% 甲酸)(B)组成。流速设定为 0.7 mL/min,梯度如下:20% B(初始条件)维持 3 分钟,4 分钟内升至 45% B;然后维持 95% B 2 分钟,最后平衡至初始条件,20% B,1 分钟。QQQ/MS 在负模式下操作。气体温度设定为 300 °C,气体流量为 12 L/min。毛细管电压设定为 5 kV.
为了检测胆汁酸,从水中回收的样品(第二部分)中取5 µL注入到Poroshell 120 EC-C8(100 mm × 2.1 mm,粒径2.7 µm;安捷伦科技)柱中,柱前保护由一根保护柱(XDB-C18,5 mm × 2.1 mm,粒径1.8 μm)提供。流动相由新鲜制备的0.2%甲酸(A)和ACN/IPA(L/L;v/v)(B)组成。流速设定为0.5 mL/min,梯度如下:30% B在2 分钟内增加到38%;保持2 分钟,然后在1.5 分钟内增加到60%,最后在2 分钟内增加到98% B(柱清洗),随后在30% B(初始条件)下进行2 分钟的柱平衡。QQQ/MS在负模式下操作。气体温度和流量分别设定为310 °C和12 L/min。毛细管电压设定为5 kV.
多胺谱分析使用第四个分馏进行:将10 μL样品注入Kinetex C18柱(150 mm × 2.1 mm,粒径2.6 µm;Phenomenex),并由一个保护C18柱(5 mm × 2.1 mm,粒径1.8 μm)保护。分析期间,柱温保持在40 °C。梯度流动相由新鲜制备的0.1% HFBA(Sigma)(A)和ACN(0.1% HFBA)(B)组成。流速设定为0.4 mL/min,梯度如下:从5%(初始条件)在7分钟内升至30% B;然后维持90% B 2分钟,最后回到初始条件(5% B,2分钟)。QQQ/MS在正模式下操作。气体温度设定为350 °C,气体流量为12 L/min。毛细管电压设定为2.5 kV。
组织样本被注入到 Zorbax Eclipse plus C18 (100 mm × 2.1 mm, 粒径 1.8 μm, Agilent) 柱中进行核苷酸和辅因子的分析,该柱由 C18 保护柱 (5 mm × 2.1 mm, 粒径 1.8 μm) 保护。在分析过程中,柱温保持在 40 °C。梯度流动相由 0.5 mM DBAA (Sigma) (A) 和 ACN (B) 组成。流速设定为 0.4 mL/min,梯度如下:10% B (初始条件) 维持 3 分钟,1 分钟内增加到 95% B,维持 2 分钟,最后平衡到初始条件,10% B,维持 1 分钟。QQQ/MS 在正负模式下均可操作。气体温度设定为 350 °C (气体流量:12 L/min)。在正模式下,毛细管电压设定为 4.5 kV,在负模式下设定为 5 kV。
在GC和UHPLC/MS中,使用了多反应监测(MRM)扫描模式进行靶向分析。峰检测和积分是使用安捷伦质谱猎人定量软件(B.10.1)进行的。
广泛针对细胞内代谢物的分析
GC/MS
一微升衍生化样品(第三分馏)被注入气相色谱仪(Agilent 7890B;安捷伦科技,德国瓦尔德布龙)与三重四极杆质谱仪(QQQ/MS;7000 C安捷伦科技,德国瓦尔德布龙)联用,配备高灵敏度电子轰击源(EI),以正模式运行。注射采用无分流模式。前进气口温度保持在250 °C,转移线和离子源温度分别为250和230 °C。隔膜清洗流量固定在3 mL/min。分流通道的清洗流量在1分钟内为80 mL/min,5分钟后气体节省模式设置为15 mL/min。氦气以1 mL/min的流速通过色谱柱(HP-5MS,30 m × 0.25 mm,内径0.25 mm,涂层J&WScientific,安捷伦科技公司)。色谱柱温度保持在60 °C 1分钟,升至210 °C(10 °C/min),然后升至230 °C(5 °C/min),最终达到325 °C(15 °C/min),并保持5分钟。碰撞气体为氮气。
假靶向分析细胞内代谢物
代谢物谱分析使用Dionex Ultimate 3000 UHPLC系统(Thermo Fisher Scientific)与Orbitrap质谱仪(q-Exactive,Thermo Fisher Scientific)联用,配备了在正负模式下均可操作的电喷雾源,并以全扫描分析模式(100–1200 m/z)获取样品。LC分离在反相(Zorbax Sb-Aq 100 × 2.1 mm × 1.8 µm粒径)上进行,流动相为:0.2%醋酸(A)和ACN(B)。柱温保持在40 °C。注入10微升水相样品(第二分馏)进行代谢物分离,梯度从2% B开始,在22分钟内增加到95% B,并维持2分钟以进行柱清洗,随后在2% B下平衡柱子4分钟。流速设定为0.3 mL/min。q-Exactive参数如下:护套气流速55 au,辅助气流速15 au,喷雾电压3.3 kV,毛细管温度300 °C,S-Lens RF水平55 V。使用醋酸钠溶液对质谱仪进行校准(专用于低质量校准)。数据最终通过Thermo XcaliburTM(版本2.2)的定量节点以伪靶向方法进行分析,使用自制的代谢物列表。
代谢组数据集的统计分析
所有目标处理的数据都通过一个专用的 R(版本 3.4)包合并和清理(@Github/Kroemerlab/GRMeta)。
RNA提取、cDNA合成、定量PCR (qPCR) 基因表达分析
从HepG2细胞、肝脏组织和附睾脂肪组织提取的总RNA分别使用RNeasy Plus Mini试剂盒(Qiagen, Hilden, Germany, #74134)和RNeasy Lipid Tissue Mini试剂盒(Qiagen, Hilden, Germany, #74804)按照制造商的说明进行提取,随后进行反转录(RT)以合成cDNA(Thermo Fisher Scientific, #11754050)。在StepOnePlus实时PCR系统(Applied Biosystems)上使用TaqMan基因表达主混合液(Applied Biosystems)进行实时定量PCR(RT qPCR)。探针(小鼠 Acbp: #Mm01286585_g1, 小鼠 Pparg: #Mm00440940_m1, 小鼠 Ppia: #Mm02342430_g1, 人类 ACBP: #Hs01554584_m1, 人类 PPARG: #Hs01115513_m1, 和人类 GAPDH: #Hs03929097_g1)购自Thermo Fisher Scientific.
RNA测序文库准备
根据制造商的说明,使用 RNeasy Plus Mini Kit 从小鼠肝脏中提取 RNA,随后进行 mRNA 测序文库准备(每个样本 1.5 μg 总 RNA;植物和动物真核生物链特异性 mRNA 和测序)。测序在 NovaSeq 6000 PE150 仪器上进行(2 × 150 bp,每个样本 4000 万条读取)。
RNA测序数据分析
伪比对和定量使用HISAT2算法(参考基因组 GRCm39) [28]。相关性分析、主成分研究和差异表达分析使用DESeq2包 [29]。差异基因表达分析使用带有Benjamini-Hochberg调整的参数Wald检验(_p_adj)。_p_adj < 0.05且log2倍数变化为±1的基因被认为是显著差异表达基因。基于基因集富集分析(GSEA)的基因本体(GO)分析,包括生物过程(BP),是在来自肝脏样本的RNA-seq数据上进行的 [30]。图表使用基于网络的生物信息学工具构建(http://www.bioinformatics.com.cn)。
染色质提取和PPARγ免疫沉淀
从小鼠肝脏中提取染色质是根据制造商的说明进行的(Diagenode, Liège, Belgium, #C01010055),随后进行超声处理和使用抗PPARγ(每次免疫沉淀10 μg,Abcam, #ab178860)和抗H3K4me3(每次免疫沉淀1 μg,Diagenode, #C01010055)抗体的免疫沉淀。简而言之,0.15 g 快速冷冻的组织被切碎并交联(1% 无甲醇的甲醛;Thermo Fisher Scientific, #28908)在室温下进行6分钟,随后在冰上用2 M甘氨酸溶液的1/10体积进行淬灭2分钟。提取的染色质经过超声处理(Diagenode, Bioruptor Pico)15个循环(30 秒“开”,30 秒“关”,位置“高”),生成平均大小约为150–300 bp的DNA片段。
ChIP-sequencing 文库准备
ChIP样本在50 μL缓冲液C(Diagenode, #C01010055)中洗脱,其中35 μL用于ChIP-sequencing(ChIP-seq)文库准备。文库使用TruSeq ChIP文库准备试剂盒(Illumina)生成,并在Illumina NovaSeq 6000上进行测序(双端,100 bp)。读取与小鼠参考基因组(mm10)使用bwa-mem 0.7.17-r1188对齐 [31]。无信息读取(多重映射、重复和低映射分数读取)使用samtools 1.3过滤 [32]。使用MACS2 2.1.1 [33]调用峰值,PPARγ ChIP-seq选项为narrow,H3K4me3标记为broad。对于每个样本,ChIP-seq根据各自的输入DNA样本进行标准化。ChIP-seq信号轨迹使用macs2生成,使用bdgcmp(和–m FE计算染色质IP样本与对照之间的倍增富集)。使用BedGraphToBigWig将文件转换为二进制格式(BigWig)。
染色质免疫沉淀定量PCR (ChIP qPCR)
ChIP样品根据制造商的说明(Diagenode, #C01010055)在50 μL中洗脱,然后稀释1:5,最后作为实时qPCR的模板(Power SYBR Green PCR Master Mix; Applied Biosystems, #4367659):DNA模板3 μL,H2O 2 μL,SYBR Green Master Mix 5 μL,[寡核苷酸] = 10 μM 2 μL.
韦恩图
RNA测序数据来自牛脂肪细胞(GSE79347)、小鼠肝细胞(GSE77625)和人类白细胞(GSE145412),这些细胞接受了常规和高脂饮食。差异表达基因通过基因表达综合数据库1进行评估(参考文献 34,35,36),使用DESeq2包进行分析 29,并基于TRRUST软件进行上游分析,使用切割阈值排除−1到+1之间的值 37。
ACBP 表达的元分析来自 GTEX 和 GEO 数据集
人类(基因型-组织表达,GTEX)、小鼠(GEO)和大鼠(GEO)基因表达数据被提取。计算了ACBP与各种感兴趣基因之间表达的Bravais–Pearson相关性(R)在GEPIA2(http://gepia2.cancer-pku.cn)上。数据使用R包ComplexHeatmaps和Circlize放入热图中。树状图由基于一个欧几里得距离矩阵的完全链接聚类组成。加权 p 值由平均R值符号和学生 t-检验的 p 值组成,零假设为(H0: µ = µ0 = 0)。
ACBP 基因启动子结合基序分析
使用基因组浏览器和ENCODE数据库对ACBP启动子进行分析,使我们能够确定可能参与_ACBP_表达调控的预测转录因子(TFs)。
方法细节
所有体外和体内实验至少重复三次 (n ≥ 3),根据我们的经验,这是获得显著结果的最佳选择,结果相似。数据以须状图(每个点代表一个生物重复)报告,包括均值 ± SEM 或热图。样本大小在图中注明。在样本超过八个 (n > 8) 的情况下,进行了正态性检验和方差齐性检验(F 或 Bartlett)。统计显著性使用未配对双尾学生 t est、未配对双尾学生 t est(Welch 校正)、Mann–Whitney 检验、单因素方差分析或双因素方差分析进行分析。当 p 值 < 0.05 时,差异被认为具有统计学意义;当 p > 0.05 时,则认为不显著 (ns)。免疫印迹密度定量以目标蛋白带与 β-肌动蛋白的比率报告(除非另有说明)。
一项生物信息学分析旨在揭示哪些转录因子(TFs)与主要代谢组织(人肝、皮下和内脏脂肪组织)中 ACBP mRNA 表达最相关,结果表明 PPARG mRNA 水平与 ACBP 的水平最相关(GEPIA2,GTEX 数据库,图 1A, B)。 ACBP 在许多其他人源组织中也与 PPARG 相关(图 1B 和图 S1A),在包括小鼠(图 1B 和图 S1B)和大鼠(图 1B 和图 S1C)的啮齿动物组织中也如此,表明物种间的保守性。这适用于人类和啮齿动物的肝脏和白色脂肪组织,以及人类的皮下脂肪组织、骨骼肌和所有组织的汇总(图 1B)。对牛脂肪细胞、小鼠肝细胞和人类白细胞的 RNA 测序(RNA-seq)数据进行了分析,以识别在正常或肥胖饮食下调节的目标基因的 TFs。只有一个单一 TF PPARγ 的目标基因在这三种细胞类型和物种的肥胖条件下始终被激活(图 1C)。因此,我们决定专注于 PPARγ。小鼠肝脏中与 PPARγ 相关的染色质的染色质免疫沉淀(ChIP)测序确认 PPARγ 结合到 Acbp 的启动子上,该区域具有真染色质标记 H3K4me3(图 1D)。此外,在人类 HepG2 肝细胞中对多个预测与 ACBP 启动子结合的 TFs(和 DNA 结合蛋白)的敲低(图 S1D)确认 PPARγ 对 ACBP mRNA 和蛋白质的基础表达是必需的(图 1E–G 和图 S1E)。
图 1: PPARγ 转录因子调节 ACBP 的表达。

A 热图表示人类肝脏、皮下白色脂肪组织 (scWAT) 和内脏白色脂肪组织 (viscWAT) 中 ACBP mRNA 与多个基因 mRNA 之间的相关性 (R) ( *p < 0.05)。B 人类 (肝脏: n = 179, 内脏 WAT: n = 355)、小鼠 (肝脏: n = 179, 附睾 WAT: n = 56) 和大鼠 (肝脏: n = 207, 附睾 WAT: n = 47) 提取物中 PPARG 和 ACBP mRNA 的相关性图。人类来源的皮下白色脂肪组织 (n = 442)、骨骼肌 (n = 304) 和所有组织的汇总 (n = 7172) 中 PPARG 和 ACBP mRNA 的相关性图。C 韦恩图表示在其供体接受高脂饮食时,牛脂肪细胞、小鼠肝细胞和人类白细胞中上调的转录因子 (TFs) 的靶标 (左)。每个数据集中 PPARγ 靶基因上调的显著性 (右)。D PPARγ 和 H3K4me3 (Acbp 启动子,小鼠肝脏, n = 3) 的染色质免疫沉淀测序 (ChIP-seq) 信号。黑线对应于每个 MACS 调用的峰值。E 在 HepG2 细胞中沉默编码人类 ACBP 的 TFs。F 在沉默无关阴性对照、ACBP 或 PPARG (siUNR, siACBP, 或 siPPARG) 后定量 ACBP 的细胞流式峰值。G 在 HepG2 细胞中沉默编码 ACBP 的各种 TFs 后的细胞流式 ACBP 蛋白水平的热图表示 (n = 3; 单因素方差分析)。对于统计分析 (A, B) p 值和 R 分别通过 Pearson 和 Spearman 相关性计算。另见图 S1。
ACBP 和 PPARγ 互相诱导
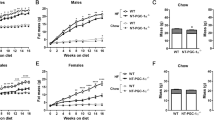
几种药理学PPARγ激动剂,包括抗糖尿病噻唑烷二酮(罗格列酮,依达列酮)以及化学上无关的工具化合物(GW1929,S26948)在人的HepG2细胞中提高了ACBP蛋白水平(图 2A)。与这些结果一致,用罗格列酮培养小鼠Hepa1–6和Hep55.1c肝癌细胞导致ACBP水平增加(图 S2A–C)。噻唑烷二酮诱导的ACBP上调通过敲低 PPARG 被逆转(图 2B),证明了该治疗的特异性。在Hep55.1 C细胞中也获得了类似的结果,其中罗格列酮诱导了PPARγ和ACBP蛋白的增加,而这一效应通过敲低 Acbp 被逆转(图 2C–E)。对小鼠进行短期(5天)每日腹腔(i.p.)注射罗格列酮的处理导致肝脏中 Pparg 和 Acbp 表达增加(图 2F)以及附睾脂肪组织(WAT)(图 S2D–F),同时血浆ACBP浓度升高(图 2G),伴随体重轻微(增加3%)但显著的变化(p = 0.002,未配对的Student t ext{-test})(图 2H)。同样,在长期(2个月)用罗格列酮处理后,小鼠表现出PPARγ、ACBP和脂肪酸合成酶(FASN)的蛋白表达增加,FASN是另一个PPARγ靶基因 \40, \41\](图 2I–L)。值得注意的是,所有这些罗格列酮诱导的变化,包括体重增加,在可诱导的全身ACBP敲除后被消除(图 2I–M)。短期(5天)用贝克沙汀(视黄醇X受体α的激动剂,RXRα,PPARγ的共激活因子,图 S3A)的处理也在肝脏中诱导了PPARγ、ACBP和FASN蛋白(图 3A, B),而RXRα抑制剂HX531则显示出相反的效果(图 3C, D)。这些体内药理学实验强调了PPARγ和ACBP之间的共同调控。
图2:PPARγ调节剂的作用依赖于ACBP功能。

A ACBP蛋白在PPARγ激动剂处理后(Ctr: 载体,Rosi: 罗格列酮,GW1929,S26948,Eda: edaglitazone)进行的细胞流式测定(n = 6)在对照(siUNR)或_ PPARG -沉默(siPPARG)的HepG2细胞中(n = 5)(B)(MFI: 相对于对照的平均荧光强度)。C 在对照(shUNR)和 Acbp -敲低(shAcbp)的Hep55.1c细胞中,处理载体或Rosi(48 h)后的PPARγ、ACBP和β-actin蛋白的代表性免疫印迹图像,密度计定量(n = 3)(D,E)。F 从接受Rosi或载体(5天)的小鼠中获得的肝脏提取物中_Pparg_和_Acbp mRNA表达测量(n = 10到13只小鼠每个条件)。G 血浆ACBP浓度(n = 7到12只小鼠每个条件),以及H 从接受Rosi或载体(5天)的小鼠中测量的体重(n = 7到8只小鼠每个条件)。I 从接受载体或Rosi(5天)的ACBP对照(ubi:Acbp WT)或ACBP敲除(ubi:Acbp KO)小鼠中FASN、PPARγ、ACBP和β-actin蛋白的肝脏代表性免疫印迹图像,密度计定量(n = 4到8只小鼠每个条件)(J–L)。M 从接受Rosi或载体(2个月)的小鼠中测量的体重(n = 4到6只小鼠每个条件)。结果以须状图显示(每个点代表一个体外生物重复或一只单独的小鼠),包括均值±SEM。对于统计分析,_p_值(表示与对照条件的统计比较)通过双尾非配对Student's _t_检验计算。对于统计分析,_p_值通过双尾非配对Student's _t_检验计算(G,H,J–L),应用Welch校正(F),单因素方差分析(A,B),或双因素方差分析(D,E,M)。MFI平均荧光强度,a.u.任意单位,kDa千道尔顿,sh短发夹,ubi普遍,ns不显著。另见图 S2。
图 3:药理学和饮食 PPARγ 操控调节 Acbp 的表达。

A 小鼠肝脏代表性免疫印迹图像,显示PPARγ、ACBP、FASN和β-actin蛋白,接受对照(载体)或RXRα激动剂Bexarotene(Bex),密度定量(n = 4到9只小鼠每组)(B)。C 小鼠肝脏代表性免疫印迹图像,显示PPARγ、ACBP、FASN和β-actin蛋白,接受对照(载体)或RXRα拮抗剂HX531药物(5天),密度定量(n = 4到8只小鼠每组)(D)。E 从接受常规饲料(RCD)或高脂饮食(HFD)的小鼠获得的肝脏提取物的定性α-PPARγ染色质免疫沉淀(ChIP)分析。ChIP PCR产物来自用PPARγ特异性抗体(α-PPARγ)沉淀的染色质样本。负同种型对照(IgG)沉淀的染色质样本中无产物。来自全身PPARγ敲除小鼠(ubi:Cre PPARγ KO)的α-PPARγ样本中无ChIP PCR产物(n = 3每组)。F ChIP实时PCR的定量分析(n = 4到8只小鼠每组)。G,H 肝脏和K,L 附睾白色脂肪组织(eWAT)Pparg_和_Acbp mRNA表达测量,来自接受RCD或HFD(6周)的小鼠(n = 7到13只小鼠每组)。I 小鼠肝脏代表性免疫印迹图像,显示FASN、PPARγ、ACBP和β-actin蛋白,接受RCD或HFD(6周),密度定量(n = 5只小鼠每组)(J)。结果以须状图显示(每个点代表一只小鼠),包括均值±SEM。对于统计分析,_p_值通过双尾非配对Student's t ext{-test}(G,H,K,L)计算,应用Welch校正(B,D,J),或双向ANOVA(F)。kDa千道尔顿,a.u.任意单位,bp碱基对,ns不显著。另见图 S3。
根据我们最近的发现 [11],血浆 ACBP 水平与短期(1 个月)高脂饮食(HFD)期间的体重增加呈正相关(图 S3B, C)。这种脂肪合成方案有利于 PPARγ 在体内与 Acbp 启动子的结合,正如通过 PPARγ 染色质免疫沉淀(ChIP)和定量逆转录酶 PCR(ChIP-qRT PCR)所示(图 3E, F 和图 S3D, E)。这种 HFD 效应伴随着肝脏中 Pparg 和 Acbp mRNA(图 3G, H)及蛋白质水平的增加(图 3I, J),以及在附睾白色脂肪组织提取物(eWAT,图 3K, L)中的增加。
总体而言,这些结果表明 ACBP 和 PPARγ 在多个组织中是共同调节的,响应于 PPARγ 激动剂、RXR 调节剂以及高脂饮食。
PPARγ去除抑制高脂饮食诱导的ACBP诱导,反之亦然
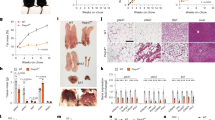
在下一步中,我们通过使用携带编码Cas9蛋白和_Pparg_特异性单导RNA(sgRNAs)的肝靶向腺相关病毒载体(AAV8)抑制了肝脏中_Pparg_的表达(图 S4A)。结果导致肝细胞特异性_Pparg_基因敲除,导致小鼠肝脏中_Pparg_ mRNA和蛋白质的整体减少,无论是接受常规饮食(RCD)的小鼠,还是接受高脂饮食(HFD)的小鼠——后者本会表现出PPARγ的上调(图 4A–C)。PPARγ的缺失减少了HFD诱导的_Acbp_肝脏mRNA和ACBP血浆蛋白的增加(图 4D, E)。这伴随着HFD诱导的体重增加的减少(图 4F),减轻了肝脏脂肪变性(图 4G, H),减少了HFD诱导的脂肪酸的局部积累(图 S4B–E),以及逆转了HFD诱导的高血糖(图 4I)。
图4:肝脏特异性PPARγ敲除导致ACBP水平降低。

A Pparg mRNA 表达 (n = 5 到 15 只小鼠每个条件) 和 B, C 蛋白水平测量 (n = 4 到 7 只小鼠每个条件) 在从对照组 (PPARγ WT) 或肝脏特异性 CRISPR/Cas9 介导的 PPARγ 敲低小鼠 (PPARγ KD) 中获得的肝脏提取物中,这些小鼠接受常规饮食 (RCD) 或高脂饮食 (HFD)。对于统计分析 (C) p 值是通过比较 HFD 组与每个遗传背景 (PPARγ WT, PPARγ KD) 对应的 RCD 组计算的。D Acbp mRNA 表达 (n = 5 到 13 只小鼠每个条件), E 血浆 ACBP 浓度 (n = 5 到 15 只小鼠每个条件), 和 F 在对照或 PPARγ KD 小鼠中测量的体重增加 (HFD, 2 个月) (n = 9 到 10 只小鼠每个条件)。G 从接受 RCD 或 HFD 的小鼠中获得的对照或 PPARγ KD 肝脏的代表性苏木精-伊红 (HE) 图像 (n = 15 到 25 只小鼠每个条件),肝脂肪变性评分量化 (条: 50 μm, ND 未检测到) (H)。I 从接受 RCD 或 HFD 的对照或肝脏 PPARγ KD 小鼠中测得的禁食血糖浓度 (n = 9 到 10 只小鼠每个条件)。结果以须条图显示 (每个点代表一只小鼠),包括均值 ± SEM。对于统计分析 p 值是通过双尾非配对学生 t-检验 (C), 双向方差分析 (A, D–F, I), 或 Mann–Whitney 检验 (H) 计算的。kDa 千道尔顿, a.u. 任意单位, ns 无显著性, ND 未检测到。另见图 S4。
接下来,我们注射了一种中和抗-ACBP抗体(图 5A),这降低了 Acbp 在肝脏中的 mRNA 表达水平,以及在高脂饮食(HFD)背景下循环的 ACBP 蛋白(图 5B, C)。ACBP 中和还降低了肝脏、eWAT 和棕色脂肪组织(BAT)中 PPARγ 蛋白的表达,无论是在接受 RCD 的小鼠中(图 5D, E)还是在接受 HFD 的小鼠中(图 5F, G)。与我们之前的发现一致 [9],全身 ACBP 中和减少了非酒精性脂肪肝病(NAFLD)的迹象,包括局部炎症(图 S5A–C),并改善了 HFD 引起的高血糖效应(图 5H),同时增加了酮体 3-羟基丁酸(图 S5D)。值得注意的是,ACBP 中和降低了 HFD 引起的血浆瘦素水平(图 S5E)。我们还确定了特定于脂肪细胞的 ACBP 基因敲除的影响(图 S5F–H),这使其对 HFD 引起的肥胖具有抵抗力 [10]。在脂肪细胞中去除 ACBP 消除了 HFD 引起的 PPARγ 上调,无论是在附睾 WAT 还是在 BAT 中(图 5I–L)。
图5:中和或基因去除ACBP导致PPARγ减少。

A ACBP中和抗体(α-ACBP)或同型对照(Iso)通过腹腔注射在喂食常规饲料(RCD)或高脂饮食(HFD)的小鼠中施用。 B 从接受Iso或α-ACBP的RCD或HFD喂养方案的小鼠中获得的肝脏提取物中测量的_Acbp_ mRNA表达(n = 5到9只小鼠每个条件)。 C 在Iso或α-ACBP处理的小鼠中测量的血浆ACBP浓度(n = 8到10只小鼠每个条件)。 D, F 从接受Iso或α-ACBP(RCD或HFD)的小鼠中提取的肝脏、附睾白色脂肪组织(eWAT)和棕色脂肪组织(BAT)中PPARγ和β-actin蛋白的代表性免疫印迹图像,密度定量(RCD: n = 5到10只小鼠每个条件, HFD: n = 4到10只小鼠每个条件)(E, G)。 H 从接受Iso或α-ACBP(RCD或HFD)的小鼠中测量的禁食血糖浓度(n = 5到10只小鼠每个条件)。 I, J 来自脂肪细胞特异性ACBP敲除小鼠模型(adipo: Acbp KO)与对照(adipo: Acbp WT)相比的eWAT和BAT中PPARγ和β-actin蛋白的代表性免疫印迹图像,密度定量(n = 5到7只小鼠每个条件)(K, L)。结果以须状图显示(每个点代表一只小鼠),包括均值±SEM。对于统计分析,_p_值通过双尾非配对Student’s t-检验计算(C, E, G, K, L)或双向ANOVA(B, H)。a.u. 表示任意单位,ns 表示不显著,kDa 表示千道尔顿。另见图 S5。
总之,基因去除PPARγ消除了高脂饮食诱导的ACBP上调,而反之,中和或敲除ACBP则阻止了高脂饮食诱导的PPARγ上调。总的来说,这些结果确认了ACBP和PPARγ之间存在一个高脂饮食触发的正反馈循环。
GABAAR中ACBP结合域的突变赋予对高脂饮食的抗性
五聚体 GABAAR 的 γ2 亚基突变(位置 77 的苯丙氨酸被异亮氨酸替代,以下简称为 F77I)据报道会消除 ACBP 信号 [24, 42]。因此,ACBP 蛋白可以从野生型小鼠(WT)中与 GABAAR γ2 亚基免疫沉淀,但不能与携带 Gabrg2 F77I 等位基因的同源小鼠(基因型:Gabrg2__F77I/F77I)的 γ2 亚基免疫沉淀(图 6A)。Gabrg2__F77I/F77I 小鼠对重组 ACBP 蛋白给药引起的多食症具有抵抗力 [10]。因此,我们研究了 GABAAR 是否与脂肪合成相结合。高脂饮食(HFD)导致 GABAAR γ2 WT 蛋白的上调,但未导致 GABAAR γ2 F77I 蛋白在肝脏中的上调(图 6B, C)。类似地,Ob/Ob 小鼠(缺乏瘦素并在正常饮食中过量进食) [43] 与其瘦小鼠 Ob/T 相比,上调了肝脏 GABAAR γ2 蛋白以及 ACBP 和 FASN 的增加(图 S6A-C)。尽管 HFD 增加了血浆 ACBP 水平(与 Gabrg2 基因背景无关,图 6D),但 Gabrg2__F77I/F77I 小鼠的肝脏中 ACBP 和 PPARγ 蛋白水平在短期(1 个月,图 6E, F)和长期(4 个月,图 S6D, E) HFD 中均下调。与这些观察一致,HFD 引起的甘油三酯和胆固醇合成在 Gabrg2__F77I/F77I I 小鼠的肝脏中减少,表明脂肪合成受损(图 S6F–I)。值得注意的是,Gabrg2__F77I/F77I 小鼠的上述代谢效应与食欲减退无关(图 S6J)。
图6:GABAAR – ACBP 受损信号的代谢效应。

A 共同免疫沉淀 (co-IP) 描述了在高脂饮食下小鼠肝提取物中 ACBP 与 GABAAR γ2 亚基之间的物理相互作用 (n = 3 每个条件)。 B 接受常规饮食 (RCD) 或高脂饮食 (HFD) 的小鼠 GABAAR γ2 亚基和 β-actin 蛋白的肝脏代表性免疫印迹图像 (1 个月),密度定量 (n = 5 到 9 只小鼠每个条件) (C)。 D 通过 RCD 或 HFD 喂养的 WT 和 F77I 小鼠的血浆 ACBP 浓度测量 (1 个月) (n = 5 到 7 只小鼠每个条件)。 E 接受 HFD (1 个月) 的 WT 和 F77I 小鼠 PPARγ 和 ACBP 蛋白的肝脏代表性免疫印迹图像,密度定量 (n = 5 只小鼠每个条件) (F)。 G 肝脏切片的代表性 HE 图像,H WT 或 F77I 小鼠在 HFD 1 个月后的肝脂肪变性评分定量 (条:50 μm,ND 未检测到) (n = 3 到 5 只小鼠每个条件)。 I 通过 RCD 或 HFD 喂养的 WT 和 F77I 小鼠的体重测量 (n = 8 到 23 只小鼠每组)。 J 表示为面积 log2 倍增变化 (Area Log2FC) 的脂肪酸、脂质和碳水化合物血浆浓度的热图表示,来自 RCD 或 HFD 喂养的 WT 或 F77I 小鼠 (1 个月) (n = 3 到 13 只小鼠每个条件),随后是代表性脂质代谢相关代谢物的定量 (K)。 结果以须状图显示 (每个点代表一只小鼠),包括均值 ± SEM。 对于统计分析,p 值通过双尾非配对学生 t est (F, K) 计算,应用 Welch 校正 (C),双向 ANOVA (D, I),或 Mann–Whitney 检验 (H)。 kDa 千道尔顿,a.u. 任意单位,ns 无显著性,ND 未检测到。另见图 S6。
鉴于 ACBP 信号传导促进了多种与肥胖相关的特征(包括肝脂肪变性、体重增加和高脂血症) [9],我们询问中和 GABAAR—ACBP 信号传导是否足以改善肥胖。有趣的是,与其 WT 同类相比,HFD 饲喂的 Gabrg2__F77I/F77I 小鼠表现出减少的肝脂肪变性(图 6G, H),对 HFD 引起的体重增加具有抵抗力(图 6I),并且循环游离脂肪酸和胆固醇水平降低(图 6J, K)。肝脏 RNA-seq 分析显示,在 GABAAR WT 小鼠中,由 HFD 上调的几个基因(包括促进肥胖的 Lcn2 基因)未能在 Gabrg2__F77I/F77I 小鼠中诱导(图 S6K, L)。
总的来说,这些结果确认了GABAAR γ2是涉及ACBP和PPARγ的前馈电路的一个重要组成部分。
我们的数据支持存在一个三元肥胖放大循环,该循环在高脂饮食小鼠的肝脏中被激活。这涉及到 (i) 促脂肪生成和刺激食欲的因子 ACBP,(ii) ACBP 受体 GABAAR,以及 [3] 转录因子 PPARγ,它在 ACBP-GABAAR 相互作用的下游转激活 ACBP 基因。支持这一三元组的论点可以总结如下:
首先,ACBP在各种组织中受到高脂饮食(HFD)的诱导,导致其血浆水平增加。脂肪细胞特异性或全身敲除ACBP,以及其抗体介导的中和,减少了HFD诱导的脂肪合成、肝脂肪变性和高血糖 [9]。此外,中和ACBP减少了HFD对PPARγ的激活。相反,系统性注射ACBP蛋白上调肝细胞中的PPARγ,因为它刺激脂肪生成 [9]。
其次,HFD或正常饮食的过量摄入(在缺乏瘦素的 Ob/Ob 小鼠中)增加了肝脏中GABAAR的γ2亚基的表达。然而,HFD未能增加无法结合ACBP的突变GABAAR(F77I)的γ2亚基的表达。Gabrg2__F77I/F77I 小鼠在重组ACBP给药后也未能产生食欲刺激反应 [10],并且与其WT同类相比,表现出减弱的HFD诱导的肝脂肪变性,以及减少的PPARγ激活和ACBP表达。
第三,HFD 增加了 PPARγ 与小鼠肝脏中 Acbp 基因启动子的结合。在体内敲低 Pparg 会减少 HFD 引起的脂肪合成、肝脂肪变性和高血糖,因为它阻止了 HFD 引起的 ACBP 增加。相反,直接用罗格列酮刺激 PPARγ,这是一种众所周知会导致患者体重增加的噻唑烷二酮类抗糖尿病药物 [44,45,46],导致 ACBP 上调,以及 ACBP 依赖的体重增加。
上述结果是在体内获得的,得到了人类和小鼠肝细胞系的体外实验的支持,这些实验表明 PPARγ 是 ACBP 基线表达所必需的,并且罗格列酮和其他 PPARγ 激动剂通过靶向效应诱导 ACBP 表达。此外,在人类组织中获得的强相关证据支持了以下假设:PPARγ 在肥胖中被激活,并且与代谢相关组织中的 ACBP 表达密切相关。
在更具推测性的层面上,饮食引起的体重增加与古老的系统的激活有关,这一点显得颇具吸引力。从进化的角度来看,ACBP 是最古老的食欲刺激物 [8]。实际上,ACBP 是唯一由单细胞真菌分泌的蛋白质。作为对饥饿的反应,它刺激孢子形成,这是此类物种唯一可能的运动形式,使它们能够寻找新的营养资源 [16, 47, 48]。同样,PPARγ 似乎是一个对营养响应的古老转录因子,其同源物已在线虫 [49]、果蝇 [50, 51],以及包括海洋硬骨鱼 [52] 和爬行动物 [53] 等多种非哺乳动物脊椎动物中被识别出来。值得注意的是,在灵长类动物进化过程中,PPARG2 中的人类特异性单核苷酸多态性已被确立 [54],这或许标志着人类特有的肥胖倾向的发展 [55]。因此,在一个理论场景中,ACBP 和 PPARγ 对体重的调节可能在动物系统发育的某个阶段变得相互关联。
值得注意的是,酵母中没有PPARγ的同源物,其中ACBP作用于G蛋白偶联受体(GPCR)STE3,这也是一种信息素受体 [16]。在小鼠中,ACBP及其肽片段十八烷神经肽(ODN)可以作用于一种可被百日咳毒素抑制的ODN-GPCR,以及GABAAR,后者不是GPCR,而是一种配体激活的氯通道 [56]。注射到大脑中的ACBP(鞘内或注入下丘脑)具有焦虑和抑食作用,这些作用可以通过ODN-GPCR抑制来阻断 [57, 58],与外周(静脉或腹腔)给药的ACBP介导的促食作用形成对比 [10]。由于全身失活ACBP和GABAAR突变具有与外周注射的抗ACBP抗体相似的代谢效应,因此外周ACBP效应似乎主导其中枢神经影响。
在这种情况下,需要注意的是,GABA,GABAAR的天然配体,已被认为与神经系统外的肥胖途径有关。因此,由GABA转氨酶催化的GABA的肝脏合成在肥胖小鼠和人群中上调,其抑制改善了小鼠的过度进食和糖尿病 [59]。GABA在肥胖小鼠的肩胛间棕色脂肪组织中也增加,且其与饮用水的口服补充足以在高脂饮食的背景下加重葡萄糖不耐受 [60]。由于GABA无法穿过血脑屏障 [61],因此这些效应应由外周(即非大脑)GABA受体介导。尽管如此,涉及GABAAR亚基的组织和细胞类型特异性切除的额外实验是必要的,以正式确定中枢或外周GABAAR信号是否决定GABA和ACBP在代谢调节中的作用。
尽管存在上述不确定性,但我们在此描述的驱动肥胖的三方前馈环似乎为治疗干预提供了几个药理学靶点。首先,可以设想对ACBP进行主动或被动免疫,以诱导中和抗ACBP自身抗体,或分别注射重组抗ACBP抗体的皮下贮存。这后一种技术已成功用于向有骨质疏松风险的女性施用抗RANKL抗体(denosumab) [62],表明其在临床环境中中和有害细胞因子的实用性。其次,可以启动努力识别小分子(理想情况下不跨越血脑屏障,但可口服)以阻断ACBP与GABAAR的γ2亚单位的相互作用。这种方法的优势在于它涉及对经典(“可药物化”)受体-配体相互作用的竞争性破坏。最后,在第三个层面上,可以尝试开发PPARγ拮抗剂。支持这一可能性的是,似乎人类PPARγ2(P12A)中的功能丧失多态性与体重指数和血糖水平降低相关 [63]。药理学PPARγ拮抗剂已在临床前模型中显示出抗肥胖和抗糖尿病活性 [64]。然而,特定阻断转录因子活性的小分子的识别 notoriously 是非常困难的 [65]。
总之,我们目前的工作识别了三种分子,(i) 致肥因子 ACBP,(ii) ACBP 受体 GABAAR,以及 (iii) ACBP 转激活转录因子 PPARγ,作为一个恶性放大循环的元素,这可能有助于肥胖及其合并症的发病机制。未来的研究将阐明这个前馈循环如何与其他致肥机制交叉,包括神经精神、内分泌、代谢、炎症、免疫和微生物回路,这些机制决定了过度体重增加的几乎不可逆转的特性。
本研究中报告的RNA-seq数据可在基因表达综合数据库(GEO)中获取,访问号为:GSE191004.
- 本瑟姆 J, 迪切萨雷 M, 比拉诺 V, 比克斯比 H, 周 B, 史蒂文斯 GA, 等. 1975年至2016年全球体重指数、体重不足、超重和肥胖的趋势:对128.9百万儿童、青少年和成人的2416项基于人群的测量研究的汇总分析. 柳叶刀. 2017;390:2627–42.
[文章](https://doi.org/10.1016%2FS0140-6736%2817%2932129-3) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Worldwide%20trends%20in%20body-mass%20index%2C%20underweight%2C%20overweight%2C%20and%20obesity%20from%201975%20to%202016%3A%20a%20pooled%20analysis%20of%202416%20population-based%20measurement%20studies%20in%20128%C2%B79%20million%20children%2C%20adolescents%2C%20and%20adults&journal=Lancet&doi=10.1016%2FS0140-6736%2817%2932129-3&volume=390&pages=2627-42&publication_year=2017&author=Bentham%2CJ&author=Cesare%2CM&author=Bilano%2CV&author=Bixby%2CH&author=Zhou%2CB&author=Stevens%2CGA)
- Swinburn BA, Kraak VI, Allender S, Atkins VJ, Baker PI, Bogard JR, 等. 肥胖、营养不良和气候变化的全球综合症:柳叶刀委员会报告. 柳叶刀. 2019;393:791–846.
[文章](https://doi.org/10.1016%2FS0140-6736%2818%2932822-8) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=30700377) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=The%20global%20syndemic%20of%20obesity%2C%20undernutrition%2C%20and%20climate%20change%3A%20the%20Lancet%20Commission%20report&journal=Lancet&doi=10.1016%2FS0140-6736%2818%2932822-8&volume=393&pages=791-846&publication_year=2019&author=Swinburn%2CBA&author=Kraak%2CVI&author=Allender%2CS&author=Atkins%2CVJ&author=Baker%2CPI&author=Bogard%2CJR)
- De Araujo IE, Schatzker M, Small DM. 重新思考食物奖励。心理学年鉴。2020;71:24.1–24.26.
[文章](https://doi.org/10.1146%2Fannurev-psych-122216-011643) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Rethinking%20food%20reward&journal=Annu%20Rev%20Psychol&doi=10.1146%2Fannurev-psych-122216-011643&volume=71&pages=24.1-24.26&publication_year=2020&author=Araujo%2CIE&author=Schatzker%2CM&author=Small%2CDM)
- Arpana Gupta VO. 肥胖和食物成瘾中的脑-肠-微生物组相互作用。生理行为。2017;176:100–6.
[谷歌学术](http://scholar.google.com/scholar_lookup?&title=Brain%E2%80%93gut%E2%80%93microbiome%20interactions%20in%20obesity%20and%20food%20addiction&journal=Physiol%20Behav&volume=176&pages=100-6&publication_year=2017&author=Arpana%20Gupta%2CVO)
- O’Brien CJO, Haberman ER, Domingos AI. 三个系统的故事:朝着肥胖的神经免疫内分泌模型迈进。细胞与发育生物学年鉴。2021;37:549–73.
[文章](https://doi.org/10.1146%2Fannurev-cellbio-120319-114106) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=34613819) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=A%20tale%20of%20three%20systems%3A%20toward%20a%20neuroimmunoendocrine%20model%20of%20obesity&journal=Annu%20Rev%20Cell%20Dev%20Biol&doi=10.1146%2Fannurev-cellbio-120319-114106&volume=37&pages=549-73&publication_year=2021&author=O%E2%80%99Brien%2CCJO&author=Haberman%2CER&author=Domingos%2CAI)
- Müller TD, Nogueiras R, Andermann ML, Andrews ZB, Anker SD, Argente J, 等. 生长激素释放肽. 分子代谢. 2015;4:437–60.
[文章](https://doi.org/10.1016%2Fj.molmet.2015.03.005) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=26042199) [PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4443295) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Ghrelin&journal=Mol%20Metab&doi=10.1016%2Fj.molmet.2015.03.005&volume=4&pages=437-60&publication_year=2015&author=M%C3%BCller%2CTD&author=Nogueiras%2CR&author=Andermann%2CML&author=Andrews%2CZB&author=Anker%2CSD&author=Argente%2CJ)
- Myers MG, Cowley MA, Münzberg H. 胰岛素作用机制及胰岛素抵抗。年复习生理学。2008;70:537–56.
[文章](https://doi.org/10.1146%2Fannurev.physiol.70.113006.100707) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BD1cXkt1eqt7k%3D) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=17937601) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Mechanisms%20of%20leptin%20action%20and%20leptin%20resistance&journal=Annu%20Rev%20Physiol&doi=10.1146%2Fannurev.physiol.70.113006.100707&volume=70&pages=537-56&publication_year=2008&author=Myers%2CMG&author=Cowley%2CMA&author=M%C3%BCnzberg%2CH)
- Montégut L, Lopez-Otin C, Magnan C, Kroemer G. 肥胖症中食欲控制的旧悖论与新机遇。内分泌代谢趋势。2021;32:264–94.
[文章](https://doi.org/10.1016%2Fj.tem.2021.02.005) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=33707095) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Old%20paradoxes%20and%20new%20opportunities%20for%20appetite%20control%20in%20obesity&journal=Trends%20Endocrinol%20Metab&doi=10.1016%2Fj.tem.2021.02.005&volume=32&pages=264-94&publication_year=2021&author=Mont%C3%A9gut%2CL&author=Lopez-Otin%2CC&author=Magnan%2CC&author=Kroemer%2CG)
- Bravo-San Pedro JM, Sica V, Martins I, Pol J, Loos F, Maiuri MC, 等。酰基辅酶A结合蛋白是一种促脂肪生成因子,触发食物摄入和肥胖。细胞代谢。2019;30:754–.e9.
[文章](https://doi.org/10.1016%2Fj.cmet.2019.07.010) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC1MXhs1SitLnN) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=31422903) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Acyl-CoA-binding%20protein%20is%20a%20lipogenic%20factor%20that%20triggers%20food%20intake%20and%20obesity&journal=Cell%20Metab&doi=10.1016%2Fj.cmet.2019.07.010&volume=30&pages=754-.e9&publication_year=2019&author=Bravo-San%20Pedro%2CJM&author=Sica%2CV&author=Martins%2CI&author=Pol%2CJ&author=Loos%2CF&author=Maiuri%2CMC)
- Joseph A, Moriceau S, Sica V, Anagnostopoulos G, Pol J, Martins I, 等. 酰基辅酶A结合蛋白(ACBP)/地西泮结合抑制剂(DBI)的代谢和精神影响. 细胞死亡与疾病. 2020;11.
- Joseph A, Chen H, Anagnostopoulos G, Montégut L, Lafarge A, Motiño O, 等. 酰基辅酶A结合蛋白(ACBP)/地西泮结合抑制剂(DBI)对身体质量指数的影响. 细胞死亡疾病. 2021;12.
- Færgeman NJ, Wadum M, Feddersen S, Burton M, Kragelund BB, Knudsen J. 酰基辅酶A结合蛋白;结构和功能在2000百万年前的保守性。分子细胞生物化学。2007;299:55–65.
[文章](https://link.springer.com/doi/10.1007/s11010-005-9040-3) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=17013545) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Acyl-CoA%20binding%20proteins%3B%20structural%20and%20functional%20conservation%20over%202000%20MYA&journal=Mol%20Cell%20Biochem&doi=10.1007%2Fs11010-005-9040-3&volume=299&pages=55-65&publication_year=2007&author=F%C3%A6rgeman%2CNJ&author=Wadum%2CM&author=Feddersen%2CS&author=Burton%2CM&author=Kragelund%2CBB&author=Knudsen%2CJ)
- Alquier T, Christian-Hinman CA, Alfonso J, Færgeman NJ. 从苯二氮卓类药物到脂肪酸及更远的地方:重新审视ACBP/DBI的作用。趋势内分泌代谢。2021;32:890–903.
[文章](https://doi.org/10.1016%2Fj.tem.2021.08.009) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB3MXitVemtLjF) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=34565656) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=From%20benzodiazepines%20to%20fatty%20acids%20and%20beyond%3A%20revisiting%20the%20role%20of%20ACBP%2FDBI&journal=Trends%20Endocrinol%20Metab&doi=10.1016%2Fj.tem.2021.08.009&volume=32&pages=890-903&publication_year=2021&author=Alquier%2CT&author=Christian-Hinman%2CCA&author=Alfonso%2CJ&author=F%C3%A6rgeman%2CNJ)
- Loomis WF, Behrens MM, Williams ME, Anjard C. 孕烯醇硫酸盐和皮质醇诱导星形胶质细胞分泌酰基辅酶A结合蛋白及其转化为内苯二氮卓类物质。生物化学杂志。2010;285:21359–65.
[文章](https://doi.org/10.1074%2Fjbc.M110.105858) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC3cXos1WrsbY%3D) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=20452969) [PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2898429) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Pregnenolone%20sulfate%20and%20cortisol%20induce%20secretion%20of%20Acyl-CoA-binding%20protein%20and%20its%20conversion%20into%20endozepines%20from%20astrocytes&journal=J%20Biol%20Chem&doi=10.1074%2Fjbc.M110.105858&volume=285&pages=21359-65&publication_year=2010&author=Loomis%2CWF&author=Behrens%2CMM&author=Williams%2CME&author=Anjard%2CC)
- Ryuda M, Tsuzuki S, Matsumoto H, Oda Y, Tanimura T, Hayakawa Y. 识别一个新基因,厌食症,通过胰岛素信号调节果蝇的进食活动。 J Biol Chem. 2011;286:38417–26.
[文章](https://doi.org/10.1074%2Fjbc.M111.267344) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC3MXhtlyktbbL) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=21917925) [PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3207462) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Identification%20of%20a%20novel%20gene%2C%20Anorexia%2C%20regulating%20feeding%20activity%20via%20insulin%20signaling%20in%20Drosophila%20melanogaster&journal=J%20Biol%20Chem&doi=10.1074%2Fjbc.M111.267344&volume=286&pages=38417-26&publication_year=2011&author=Ryuda%2CM&author=Tsuzuki%2CS&author=Matsumoto%2CH&author=Oda%2CY&author=Tanimura%2CT&author=Hayakawa%2CY)
- Charmpilas N, Ruckenstuhl C, Sica V, Büttner S, Habernig L, Dichtinger S, 等. 酰基辅酶A结合蛋白 (ACBP): 一种系统发育上保守的食欲刺激物. 细胞死亡与疾病. 2020;11.
- Bautista W, Perez-Alvarez V, Burczynski F, Raouf A, Klonisch T, Minuk G. 肝肿瘤和非肿瘤干细胞中的膜电位差异和GABAA受体表达。加拿大学报生理药理学。2014;92:85–91.
[文章](https://doi.org/10.1139%2Fcjpp-2013-0226) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC3sXhvFGrtrjF) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Membrane%20potential%20differences%20and%20GABAA%20receptor%20expression%20in%20hepatic%20tumor%20and%20non-tumor%20stem%20cells&journal=Can%20J%20Physiol%20Pharm&doi=10.1139%2Fcjpp-2013-0226&volume=92&pages=85-91&publication_year=2014&author=Bautista%2CW&author=Perez-Alvarez%2CV&author=Burczynski%2CF&author=Raouf%2CA&author=Klonisch%2CT&author=Minuk%2CG)
- Wang S, Xiang YY, Zhu J, Yi F, Li J, Liu C, 等. 肝脏GABA信号在大鼠急性肝损伤中的保护作用. Am J Physiol Gastrointest Liver Physiol. 2017;312:G208–18.
[文章](https://doi.org/10.1152%2Fajpgi.00344.2016) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=27979827) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Protective%20roles%20of%20hepatic%20GABA%20signaling%20in%20acute%20liver%20injury%20of%20rats&journal=Am%20J%20Physiol%20Gastrointest%20Liver%20Physiol&doi=10.1152%2Fajpgi.00344.2016&volume=312&pages=G208-18&publication_year=2017&author=Wang%2CS&author=Xiang%2CYY&author=Zhu%2CJ&author=Yi%2CF&author=Li%2CJ&author=Liu%2CC)
- Bu J, Huang S, Wang J, Xia T, Liu H, You Y, 等. GABAA 受体通过调节小鼠中的巨噬细胞影响压力超负荷引起的心力衰竭. Front Immunol. 2021;12:1–12.
[谷歌学术](http://scholar.google.com/scholar_lookup?&title=The%20GABAA%20receptor%20influences%20pressure%20overload-induced%20heart%20failure%20by%20modulating%20macrophages%20in%20mice&journal=Front%20Immunol&volume=12&pages=1-12&publication_year=2021&author=Bu%2CJ&author=Huang%2CS&author=Wang%2CJ&author=Xia%2CT&author=Liu%2CH&author=You%2CY)
- Bravo-San Pedro JM, Sica V, Madeo F, Kroemer G. 酰基辅酶A结合蛋白 (ACBP): 神秘的“饥饿因子”,将自噬与食物摄入联系起来。细胞压力。2019;3:312–8.
[文章](https://doi.org/10.15698%2Fcst2019.10.200) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Acyl-CoA-binding%20protein%20%28ACBP%29%3A%20the%20elusive%20%E2%80%9Chunger%20factor%E2%80%9D%20linking%20autophagy%20to%20food%20intake&journal=Cell%20Stress.&doi=10.15698%2Fcst2019.10.200&volume=3&pages=312-8&publication_year=2019&author=Bravo-San%20Pedro%2CJM&author=Sica%2CV&author=Madeo%2CF&author=Kroemer%2CG)
- Clavier T, Tonon MC, Foutel A, Besnier E, Lefevre-Scelles A, Morin F, 等. 在全身炎症期间内源性苯二氮卓受体配体内啡肽的血浆水平增加:一项前瞻性观察研究. Crit Care. 2014;18:1–8.
[文章](https://link.springer.com/doi/10.1186/s13054-014-0633-7) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Increased%20plasma%20levels%20of%20endozepines%2C%20endogenous%20ligands%20of%20benzodiazepine%20receptors%2C%20during%20systemic%20inflammation%3A%20a%20prospective%20observational%20study&journal=Crit%20Care&doi=10.1186%2Fs13054-014-0633-7&volume=18&pages=1-8&publication_year=2014&author=Clavier%2CT&author=Tonon%2CMC&author=Foutel%2CA&author=Besnier%2CE&author=Lefevre-Scelles%2CA&author=Morin%2CF)
- Conti E, Tremolizzo L, Bomba M, Uccellini O, Rossi MS, Raggi ME, 等。青少年厌食症患者空腹血浆地西泮结合抑制剂水平降低。国际饮食失调杂志。2013;46:626–9.
[文章](https://doi.org/10.1002%2Feat.22129) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=23625555) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Reduced%20fasting%20plasma%20levels%20of%20diazepam-binding%20inhibitor%20in%20adolescents%20with%20anorexia%20nervosa&journal=Int%20J%20Eat%20Disord&doi=10.1002%2Feat.22129&volume=46&pages=626-9&publication_year=2013&author=Conti%2CE&author=Tremolizzo%2CL&author=Bomba%2CM&author=Uccellini%2CO&author=Rossi%2CMS&author=Raggi%2CME)
- Li S, Joseph A, Martins I, Kroemer G. 在禁食和肥胖受试者中,食欲刺激物 acbp/dbi 的血浆水平升高。Cell Stress. 2021;5:89–98.
[文章](https://doi.org/10.15698%2Fcst2021.07.252) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB3MXit1ahurzO) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=34308254) [PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC8283301) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Elevated%20plasma%20levels%20of%20the%20appetite-stimulator%20acbp%2Fdbi%20in%20fasting%20and%20obese%20subjects&journal=Cell%20Stress&doi=10.15698%2Fcst2021.07.252&volume=5&pages=89-98&publication_year=2021&author=Li%2CS&author=Joseph%2CA&author=Martins%2CI&author=Kroemer%2CG)
- Wulff P, Goetz T, Leppä E, Linden AM, Renzi M, Swinny JD, 等. 从突触到行为:用工程化的GABAA受体快速调节特定神经元类型. Neuroforum. 2007;13:134–6.
[文章](https://doi.org/10.1515%2Fnf-2007-0405) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=From%20synapse%20to%20behavior%3A%20Rapid%20modulation%20of%20defined%20neuronal%20types%20with%20engineered%20GABAA%20receptors&journal=Neuroforum&doi=10.1515%2Fnf-2007-0405&volume=13&pages=134-6&publication_year=2007&author=Wulff%2CP&author=Goetz%2CT&author=Lepp%C3%A4%2CE&author=Linden%2CAM&author=Renzi%2CM&author=Swinny%2CJD)
- Ran FA, Cong L, Yan WX, Scott DA, Gootenberg JS, Kriz AJ, 等. 使用金黄色葡萄球菌 Cas9 的体内基因组编辑. 自然 2015;520:186–91.
[文章](https://doi.org/10.1038%2Fnature14299) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC2MXmt1Sms7o%3D) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=25830891) [PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4393360) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=In%20vivo%20genome%20editing%20using%20Staphylococcus%20aureus%20Cas9&journal=Nature&doi=10.1038%2Fnature14299&volume=520&pages=186-91&publication_year=2015&author=Ran%2CFA&author=Cong%2CL&author=Yan%2CWX&author=Scott%2CDA&author=Gootenberg%2CJS&author=Kriz%2CAJ)
- Mendler MH, Kanel G, Govindarajan S. 提出一种非酒精性脂肪肝病的组织学评分和分级系统。Liver Int. 2005;25:294–304.
[文章](https://doi.org/10.1111%2Fj.1478-3231.2005.01052.x) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=15780053) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Proposal%20for%20a%20histological%20scoring%20and%20grading%20system%20for%20non-alcoholic%20fatty%20liver%20disease&journal=Liver%20Int&doi=10.1111%2Fj.1478-3231.2005.01052.x&volume=25&pages=294-304&publication_year=2005&author=Mendler%2CMH&author=Kanel%2CG&author=Govindarajan%2CS)
- Grajeda-lglesias C, Durand S, Daillére R, Iribarren K, Lemaitre F, Derosa L, 等。口服给药Akkermansia muciniphila提高全身抗衰老和抗癌代谢物。衰老。2021;13:6375–405.
[文章](https://doi.org/10.18632%2Faging.202739) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Oral%20administration%20of%20Akkermansia%20muciniphila%20elevates%20systemic%20antiaging%20and%20anticancer%20metabolites&journal=Aging&doi=10.18632%2Faging.202739&volume=13&pages=6375-405&publication_year=2021&author=Grajeda-lglesias%2CC&author=Durand%2CS&author=Daill%C3%A9re%2CR&author=Iribarren%2CK&author=Lemaitre%2CF&author=Derosa%2CL)
- Kim D, Paggi JM, Park C, Bennett C, Salzberg SL. 基于图的基因组比对和基因分型,使用HISAT2和HISAT-genotype. Nat Biotechnol. 2019;37:907–15.
[文章](https://doi.org/10.1038%2Fs41587-019-0201-4) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC1MXhsFWqtL7O) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=31375807) [PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC7605509) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Graph-based%20genome%20alignment%20and%20genotyping%20with%20HISAT2%20and%20HISAT-genotype&journal=Nat%20Biotechnol&doi=10.1038%2Fs41587-019-0201-4&volume=37&pages=907-15&publication_year=2019&author=Kim%2CD&author=Paggi%2CJM&author=Park%2CC&author=Bennett%2CC&author=Salzberg%2CSL)
- Love MI, Huber W, Anders S. 使用 DESeq2 对 RNA-seq 数据进行折叠变化和离散度的调节估计。基因组生物学。2014;15:1–21.
[文章](https://link.springer.com/doi/10.1186/s13059-014-0550-8) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Moderated%20estimation%20of%20fold%20change%20and%20dispersion%20for%20RNA-seq%20data%20with%20DESeq2&journal=Genome%20Biol&doi=10.1186%2Fs13059-014-0550-8&volume=15&pages=1-21&publication_year=2014&author=Love%2CMI&author=Huber%2CW&author=Anders%2CS)
- Wang J, Vasaikar S, Shi Z, Greer M, Zhang B. WebGestalt 2017: 一个更全面、更强大、更灵活和互动的基因集富集分析工具包。核酸研究。2017;45:W130–7.
[文章](https://doi.org/10.1093%2Fnar%2Fgkx356) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC1cXhs1ait7w%3D) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=28472511) [PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC5570149) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=WebGestalt%202017%3A%20a%20more%20comprehensive%2C%20powerful%2C%20flexible%20and%20interactive%20gene%20set%20enrichment%20analysis%20toolkit&journal=Nucleic%20Acids%20Res&doi=10.1093%2Fnar%2Fgkx356&volume=45&pages=W130-7&publication_year=2017&author=Wang%2CJ&author=Vasaikar%2CS&author=Shi%2CZ&author=Greer%2CM&author=Zhang%2CB)
- Li H, Durbin R. 使用Burrows-Wheeler变换进行快速准确的短序列比对。生物信息学。2009;25:1754–60.
[文章](https://doi.org/10.1093%2Fbioinformatics%2Fbtp324) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BD1MXot1Cjtbo%3D) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=19451168) [PubMed 中心](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2705234) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Fast%20and%20accurate%20short%20read%20alignment%20with%20Burrows-Wheeler%20transform&journal=Bioinformatics.&doi=10.1093%2Fbioinformatics%2Fbtp324&volume=25&pages=1754-60&publication_year=2009&author=Li%2CH&author=Durbin%2CR)
- Li H, Handsaker B, Wysoker A, Fennell T, Ruan J, Homer N, 等. 序列对齐/映射格式和SAMtools. 生物信息学. 2009;25:2078–9.
[文章](https://doi.org/10.1093%2Fbioinformatics%2Fbtp352) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=19505943) [PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2723002) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=The%20sequence%20alignment%2Fmap%20format%20and%20SAMtools&journal=Bioinformatics.&doi=10.1093%2Fbioinformatics%2Fbtp352&volume=25&pages=2078-9&publication_year=2009&author=Li%2CH&author=Handsaker%2CB&author=Wysoker%2CA&author=Fennell%2CT&author=Ruan%2CJ&author=Homer%2CN)
- Zhang Y, Liu T, Meyer CA, Eeckhoute J, Johnson DS, Bernstein BE, 等. 基于模型的ChIP-Seq分析(MACS)。基因组生物学。2008;9:R137.
[文章](https://link.springer.com/doi/10.1186/gb-2008-9-9-r137) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=18798982) [PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2592715) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Model-based%20analysis%20of%20ChIP-Seq%20%28MACS%29&journal=Genome%20Biol&doi=10.1186%2Fgb-2008-9-9-r137&volume=9&publication_year=2008&author=Zhang%2CY&author=Liu%2CT&author=Meyer%2CCA&author=Eeckhoute%2CJ&author=Johnson%2CDS&author=Bernstein%2CBE)
- Soltis AR, Kennedy NJ, Xin X, Zhou F, Ficarro SB, Yap YS, 等. 由于高脂饮食引起的肝功能障碍. Cell Rep. 2017;21:3317–28.
[文章](https://doi.org/10.1016%2Fj.celrep.2017.11.059) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC2sXhvFOnsbzI) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=29241556) [PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC5734865) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Hepatic%20dysfunction%20caused%20by%20consumption%20of%20a%20high-fat%20diet&journal=Cell%20Rep&doi=10.1016%2Fj.celrep.2017.11.059&volume=21&pages=3317-28&publication_year=2017&author=Soltis%2CAR&author=Kennedy%2CNJ&author=Xin%2CX&author=Zhou%2CF&author=Ficarro%2CSB&author=Yap%2CYS)
- Wærp HKL, Waters SM, McCabe MS, Cormican P, Salte R. 先前饮食、饮食转变和怀孕对奶牛小母牛脂肪基因表达的长期影响。PLoS ONE. 2019;14:1–28.
[文章](https://doi.org/10.1371%2Fjournal.pone.0218723) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Long-term%20effects%20of%20prior%20diets%2C%20dietary%20transition%20and%20pregnancy%20on%20adipose%20gene%20expression%20in%20dairy%20heifers&journal=PLoS%20ONE&doi=10.1371%2Fjournal.pone.0218723&volume=14&pages=1-28&publication_year=2019&author=W%C3%A6rp%2CHKL&author=Waters%2CSM&author=McCabe%2CMS&author=Cormican%2CP&author=Salte%2CR)
- Paczkowska-Abdulsalam M, Niemira M, Bielska A, Szalkowska A, Raczkowska BA, Junttila S, 等. 评估肥胖和瘦体重个体中代谢综合征背后的转录组调控. Int J Mol Sci. 2020;21:1455.
[文章](https://doi.org/10.3390%2Fijms21041455) [PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC7073064) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Evaluation%20of%20transcriptomic%20regulations%20behind%20metabolic%20syndrome%20in%20obese%20and%20lean%20subjects&journal=Int%20J%20Mol%20Sci&doi=10.3390%2Fijms21041455&volume=21&publication_year=2020&author=Paczkowska-Abdulsalam%2CM&author=Niemira%2CM&author=Bielska%2CA&author=Szalkowska%2CA&author=Raczkowska%2CBA&author=Junttila%2CS)
- Han H, Cho JW, Lee S, Yun A, Kim H, Bae D, 等. TRRUST v2: 扩展的人类和小鼠转录调控相互作用参考数据库. 核酸研究. 2018;46:D380–6.
[文章](https://doi.org/10.1093%2Fnar%2Fgkx1013) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC1cXitlGisb3P) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=29087512) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=TRRUST%20v2%3A%20an%20expanded%20reference%20database%20of%20human%20and%20mouse%20transcriptional%20regulatory%20interactions&journal=Nucleic%20Acids%20Res&doi=10.1093%2Fnar%2Fgkx1013&volume=46&pages=D380-6&publication_year=2018&author=Han%2CH&author=Cho%2CJW&author=Lee%2CS&author=Yun%2CA&author=Kim%2CH&author=Bae%2CD)
- Elholm M, Bjerking G, Knudsen J, Kristiansen K, Mandrup S. 大鼠编码酰基辅酶A结合蛋白的启动子区域中的调控元件。基因。1996;173:233–8.
[文章](https://doi.org/10.1016%2F0378-1119%2896%2900213-2) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DyaK28Xlsl2gtLc%3D) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=8964505) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Regulatory%20elements%20in%20the%20promoter%20region%20of%20the%20rat%20gene%20encoding%20the%20acyl-CoA-binding%20protein&journal=Gene.&doi=10.1016%2F0378-1119%2896%2900213-2&volume=173&pages=233-8&publication_year=1996&author=Elholm%2CM&author=Bjerking%2CG&author=Knudsen%2CJ&author=Kristiansen%2CK&author=Mandrup%2CS)
- Marineau C, Aubry M, Julien JP, Rouleau GA. D22S264位点的二核苷酸重复多态性。核酸研究。1992;20:1430.
[PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=1561110) [PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC312207) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Dinucleotide%20repeat%20polymorphism%20at%20the%20D22S264%20locus&journal=Nucleic%20Acids%20Res&volume=20&publication_year=1992&author=Marineau%2CC&author=Aubry%2CM&author=Julien%2CJP&author=Rouleau%2CGA)
- Takahashi N, Kawada T, Yamamoto T, Goto T, Taimatsu A, Aoki N, 等. 过表达和核糖酶介导的转录共激活因子CREB结合蛋白和p300的靶向揭示了它们在脂肪细胞分化中的不可或缺的作用,主要通过调节过氧化物酶体增殖物激活受体γ. J Biol Chem. 2002;277:16906–12.
[文章](https://doi.org/10.1074%2Fjbc.M200585200) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BD38XjslWhtbk%3D) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=11884404) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Overexpression%20and%20ribozyme-mediated%20targeting%20of%20transcriptional%20coactivators%20CREB-binding%20protein%20and%20p300%20revealed%20their%20indispensable%20roles%20in%20adipocyte%20differentiation%20through%20the%20regulation%20of%20peroxisome%20proliferator-activated%20receptor%20%CE%B3&journal=J%20Biol%20Chem&doi=10.1074%2Fjbc.M200585200&volume=277&pages=16906-12&publication_year=2002&author=Takahashi%2CN&author=Kawada%2CT&author=Yamamoto%2CT&author=Goto%2CT&author=Taimatsu%2CA&author=Aoki%2CN)
- Salma N, Xiao H, Mueller E, Imbalzano AN. 在过氧化物酶体增殖物激活受体γ核激素受体的脂肪生成诱导过程中转录因子和SWI/SNF染色质重塑酶的时间招募。Mol Cell Biol. 2004;24:4651–63.
[文章](https://doi.org/10.1128%2FMCB.24.11.4651-4663.2004) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BD2cXksVyhsrk%3D) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=15143161) [PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC416408) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Temporal%20recruitment%20of%20transcription%20factors%20and%20SWI%2FSNF%20chromatin-remodeling%20enzymes%20during%20adipogenic%20induction%20of%20the%20peroxisome%20proliferator-activated%20receptor%20%CE%B3%20nuclear%20hormone%20receptor&journal=Mol%20Cell%20Biol&doi=10.1128%2FMCB.24.11.4651-4663.2004&volume=24&pages=4651-63&publication_year=2004&author=Salma%2CN&author=Xiao%2CH&author=Mueller%2CE&author=Imbalzano%2CAN)
- Dumitru I, Neitz A, Alfonso J, Monyer H. 地西泮结合抑制剂促进干细胞扩展,控制环境依赖的神经发生。神经元。2017;94:125–.e5.
[文章](https://doi.org/10.1016%2Fj.neuron.2017.03.003) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC2sXkvFWqt70%3D) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=28343864) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Diazepam%20binding%20inhibitor%20promotes%20stem%20cell%20expansion%20controlling%20environment-dependent%20neurogenesis&journal=Neuron&doi=10.1016%2Fj.neuron.2017.03.003&volume=94&pages=125-.e5&publication_year=2017&author=Dumitru%2CI&author=Neitz%2CA&author=Alfonso%2CJ&author=Monyer%2CH)
- Halaas JL, Gajiwala KS, Maffei M, Cohen SL, Chait BT, Rabinowitz D, 等。由肥胖基因编码的血浆蛋白的减重效果。科学。1995;269:543–6.
[文章](https://doi.org/10.1126%2Fscience.7624777) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DyaK2MXntF2ktr8%3D) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=7624777) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Weight-reducing%20effects%20of%20the%20plasma%20protein%20encoded%20by%20the%20obese%20gene&journal=Science&doi=10.1126%2Fscience.7624777&volume=269&pages=543-6&publication_year=1995&author=Halaas%2CJL&author=Gajiwala%2CKS&author=Maffei%2CM&author=Cohen%2CSL&author=Chait%2CBT&author=Rabinowitz%2CD)
- Kahn SE, Haffner SM, Heise MA, Herman WH, Holman RR, Jones NP, 等. 罗格列酮、二甲双胍或格列本脲单药治疗的血糖持久性. 新英格兰医学杂志. 2006;355:2427–43.
[文章](https://doi.org/10.1056%2FNEJMoa066224) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BD28Xht12ntrzF) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=17145742) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Glycemic%20durability%20of%20rosiglitazone%2C%20metformin%2C%20or%20glyburide%20monotherapy&journal=N%20Engl%20J%20Med&doi=10.1056%2FNEJMoa066224&volume=355&pages=2427-43&publication_year=2006&author=Kahn%2CSE&author=Haffner%2CSM&author=Heise%2CMA&author=Herman%2CWH&author=Holman%2CRR&author=Jones%2CNP)
- Kahn SE, Haffner SM, Viberti G, Herman WH, Lachin JM, Kravitz BG, 等。罗格列酮相较于格列本脲和二甲双胍在4年内显著降低C反应蛋白,尽管体重增加更明显:来自一项糖尿病结果进展试验(ADOPT)的观察。糖尿病护理。2010;33:177–83.
[文章](https://doi.org/10.2337%2Fdc09-1661) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC3cXht1Kiur8%3D) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=19808911) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Rosiglitazone%20decreases%20C-reactive%20protein%20to%20a%20greater%20extent%20relative%20to%20glyburide%20and%20metformin%20over%204%20years%20despite%20greater%20weight%20gain%3A%20observations%20from%20A%20diabetes%20outcome%20progression%20trial%20%28ADOPT%29&journal=Diabetes%20Care&doi=10.2337%2Fdc09-1661&volume=33&pages=177-83&publication_year=2010&author=Kahn%2CSE&author=Haffner%2CSM&author=Viberti%2CG&author=Herman%2CWH&author=Lachin%2CJM&author=Kravitz%2CBG)
- Bundhun PK, Janoo G, Teeluck AR, Huang F. 在治疗2型糖尿病患者时,vildagliptin与pioglitazone或rosiglitazone观察到的不良药物反应:一项随机对照试验的系统评价和荟萃分析。BMC Pharm Toxicol. 2017;18:1–10.
[谷歌学术](http://scholar.google.com/scholar_lookup?&title=Adverse%20drug%20effects%20observed%20with%20vildagliptin%20versus%20pioglitazone%20or%20rosiglitazone%20in%20the%20treatment%20of%20patients%20with%20type%202%20diabetes%20mellitus%3A%20a%20systematic%20review%20and%20meta-analysis%20of%20randomized%20controlled%20trials&journal=BMC%20Pharm%20Toxicol&volume=18&pages=1-10&publication_year=2017&author=Bundhun%2CPK&author=Janoo%2CG&author=Teeluck%2CAR&author=Huang%2CF)
- Alvarez-Curto E, Saran S, Meima M, Zobel J, Scott C, Schaap P. 腺苷酸酰化酶 G 诱导 Dictyostelium slugs 中的前孢子分化的 cAMP 产生。 发展。 2007;134:959–66.
[文章](https://doi.org/10.1242%2Fdev.02775) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BD2sXkt1Ogsbw%3D) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=17267449) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=cAMP%20production%20by%20adenylyl%20cyclase%20G%20induces%20prespore%20differentiation%20in%20Dictyostelium%20slugs&journal=Development.&doi=10.1242%2Fdev.02775&volume=134&pages=959-66&publication_year=2007&author=Alvarez-Curto%2CE&author=Saran%2CS&author=Meima%2CM&author=Zobel%2CJ&author=Scott%2CC&author=Schaap%2CP)
- Manjithaya R, Anjard C, Loomis WF, Subramani S. Pichia pastoris Acb1 的非常规分泌依赖于 GRASP 蛋白、过氧化物酶体功能和自噬体形成。J Cell Biol. 2010;188:537–46.
[文章](https://doi.org/10.1083%2Fjcb.200911149) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC3cXislWgsrg%3D) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=20156962) [PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2828923) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Unconventional%20secretion%20of%20Pichia%20pastoris%20Acb1%20is%20dependent%20on%20GRASP%20protein%2C%20peroxisomal%20functions%2C%20and%20autophagosome%20formation&journal=J%20Cell%20Biol&doi=10.1083%2Fjcb.200911149&volume=188&pages=537-46&publication_year=2010&author=Manjithaya%2CR&author=Anjard%2CC&author=Loomis%2CWF&author=Subramani%2CS)
- Van Gilst MR, Hadjivassiliou H, Jolly A, Yamamoto KR. 核激素受体 NHR-49 控制 C. elegans 中的脂肪消耗和脂肪酸组成。PLoS Biol. 2005;3:0301–12.
[文章](https://doi.org/10.1371%2Fjournal.pbio.0030053) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Nuclear%20hormone%20receptor%20NHR-49%20controls%20fat%20consumption%20and%20fatty%20acid%20composition%20in%20C.%20elegans&journal=PLoS%20Biol&doi=10.1371%2Fjournal.pbio.0030053&volume=3&pages=0301-12&publication_year=2005&author=Gilst%2CMR&author=Hadjivassiliou%2CH&author=Jolly%2CA&author=Yamamoto%2CKR)
- Joardar A, Menzl J, Podolsky TC, Manzo E, Estes PS, Ashford S, 等。PPAR gamma 激活在基于 TDP-43 的果蝇 ALS 模型中具有神经保护作用。Hum Mol Genet. 2015;24:1741–54.
[文章](https://doi.org/10.1093%2Fhmg%2Fddu587) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC2MXhsVylsbjJ) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=25432537) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=PPAR%20gamma%20activation%20is%20neuroprotective%20in%20a%20Drosophila%20model%20of%20ALS%20based%20on%20TDP-43&journal=Hum%20Mol%20Genet&doi=10.1093%2Fhmg%2Fddu587&volume=24&pages=1741-54&publication_year=2015&author=Joardar%2CA&author=Menzl%2CJ&author=Podolsky%2CTC&author=Manzo%2CE&author=Estes%2CPS&author=Ashford%2CS)
- Zipper L, Jassmann D, Burgmer S, Görlich B, Reiff T. 蜕皮激素类固醇激素通过PPARγ同源物Eip75B在果蝇中远程控制肠道干细胞命运决策。Elife. 2020;9:e55795.
[文章](https://doi.org/10.7554%2FeLife.55795) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=32773037) [PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC7440922) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Ecdysone%20steroid%20hormone%20remote%20controls%20intestinal%20stem%20cell%20fate%20decisions%20via%20the%20PPAR%CE%B3-homolog%20Eip75B%20in%20Drosophila&journal=Elife&doi=10.7554%2FeLife.55795&volume=9&publication_year=2020&author=Zipper%2CL&author=Jassmann%2CD&author=Burgmer%2CS&author=G%C3%B6rlich%2CB&author=Reiff%2CT)
- Leaver MJ, Boukouvala E, Antonopoulou E, Diez A, Favre-Krey L, Tariq Ezaz M, 等. 来自两种海洋鱼类的三种过氧化物酶增殖物激活受体亚型. 内分泌学. 2005;146:3150–62.
[文章](https://doi.org/10.1210%2Fen.2004-1638) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BD2MXlslGhtrk%3D) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=15790725) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Three%20peroxisome%20proliferator-activated%20receptor%20isotypes%20from%20each%20of%20two%20species%20of%20marine%20fish&journal=Endocrinology.&doi=10.1210%2Fen.2004-1638&volume=146&pages=3150-62&publication_year=2005&author=Leaver%2CMJ&author=Boukouvala%2CE&author=Antonopoulou%2CE&author=Diez%2CA&author=Favre-Krey%2CL&author=Tariq%20Ezaz%2CM)
- Kato K, Oka Y, Park MK. 在爬行动物豹纹壁虎 (Eublepharis macularius) 中识别和表达分析过氧化物酶体增殖物激活受体 cDNA。动物科学。2008;25:492–502.
[文章](https://doi.org/10.2108%2Fzsj.25.492) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BD1cXhtF2itLfO) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Identification%20and%20expression%20analysis%20of%20peroxisome%20proliferator-activated%20receptors%20cDNA%20in%20a%20reptile%2C%20the%20leopard%20gecko%20%28Eublepharis%20macularius%29&journal=Zool%20Sci&doi=10.2108%2Fzsj.25.492&volume=25&pages=492-502&publication_year=2008&author=Kato%2CK&author=Oka%2CY&author=Park%2CMK)
- 竹中 A, 中村 S, 三永 F, 井上-村山 M, 宇土野 T, 苏里约布罗托 B. 在灵长类动物进化过程中,与肥胖基因相关的人类特异性 SNP,肾上腺素能受体 beta2 (ADRB2)、beta3 (ADRB3) 和 PPAR γ2 (PPARG)。PLoS ONE. 2012;7:e43461.
[文章](https://doi.org/10.1371%2Fjournal.pone.0043461) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=22937051) [PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3427335) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Human-specific%20SNP%20in%20obesity%20genes%2C%20adrenergic%20receptor%20beta2%20%28ADRB2%29%2C%20beta3%20%28ADRB3%29%2C%20and%20PPAR%20%CE%B32%20%28PPARG%29%2C%20during%20primate%20evolution&journal=PLoS%20ONE&doi=10.1371%2Fjournal.pone.0043461&volume=7&publication_year=2012&author=Takenaka%2CA&author=Nakamura%2CS&author=Mitsunaga%2CF&author=Inoue-Murayama%2CM&author=Udono%2CT&author=Suryobroto%2CB)
- Jeninga EH, Gurnell M, Kalkhoven E. 转录因子抑制:经验教训与新兴靶点。内分泌代谢趋势。2009;20:380–7.
[文章](https://doi.org/10.1016%2Fj.tem.2009.04.005) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BD1MXht1Wnu7nE) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=19748282) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Transcription%20factor%20inhibition%3A%20lessons%20learned%20and%20emerging%20targets&journal=Trends%20Endocrinol%20Metab&doi=10.1016%2Fj.tem.2009.04.005&volume=20&pages=380-7&publication_year=2009&author=Jeninga%2CEH&author=Gurnell%2CM&author=Kalkhoven%2CE)
- Castellano D, Shepard RD, Lu W. 在一个“旧”受体中寻找新奇:我们对GABAARs的理解及其在受体药理学中的意义的最新进展。前沿神经科学。2021;14:1–15.
[文章](https://doi.org/10.3389%2Ffnins.2020.616298) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Looking%20for%20novelty%20in%20an%20%E2%80%9Cold%E2%80%9D%20receptor%3A%20recent%20advances%20toward%20our%20understanding%20of%20GABAARs%20and%20their%20implications%20in%20receptor%20pharmacology&journal=Front%20Neurosci&doi=10.3389%2Ffnins.2020.616298&volume=14&pages=1-15&publication_year=2021&author=Castellano%2CD&author=Shepard%2CRD&author=Lu%2CW)
- Changhan Lee VL. 有和没有热量限制的饮食限制对健康老龄化的影响。老龄化研究评论。2016;39:15–28.
[谷歌学术](http://scholar.google.com/scholar_lookup?&title=Dietary%20restriction%20with%20and%20without%20caloric%20restriction%20for%20healthy%20aging&journal=Ageing%20Res%20Rev&volume=39&pages=15-28&publication_year=2016&author=Changhan%20Lee%2CVL)
- Bouyakdan K, Martin H, Liénard F, Budry L, Taib B, Rodaros D, 等。神经胶质传递物质 ACBP 通过黑色素皮质素系统控制进食和能量稳态。J Clin Invest. 2019;129:2417–30.
[文章](https://doi.org/10.1172%2FJCI123454) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=30938715) [PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC6546475) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=The%20gliotransmitter%20ACBP%20controls%20feeding%20and%20energy%20homeostasis%20via%20the%20melanocortin%20system&journal=J%20Clin%20Invest&doi=10.1172%2FJCI123454&volume=129&pages=2417-30&publication_year=2019&author=Bouyakdan%2CK&author=Martin%2CH&author=Li%C3%A9nard%2CF&author=Budry%2CL&author=Taib%2CB&author=Rodaros%2CD)
- Geisler CE, Ghimire S, Bruggink SM, Miller KE, Weninger SN, Kronenfeld JM, 等. 肝脏GABA在肥胖的代谢功能障碍和多食症中的关键作用. Cell Rep. 2021;35:109301.
[文章](https://doi.org/10.1016%2Fj.celrep.2021.109301) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB3MXhsVKlsbbF) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=34192532) [PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC8851954) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=A%20critical%20role%20of%20hepatic%20GABA%20in%20the%20metabolic%20dysfunction%20and%20hyperphagia%20of%20obesity&journal=Cell%20Rep&doi=10.1016%2Fj.celrep.2021.109301&volume=35&publication_year=2021&author=Geisler%2CCE&author=Ghimire%2CS&author=Bruggink%2CSM&author=Miller%2CKE&author=Weninger%2CSN&author=Kronenfeld%2CJM)
- Ikegami R, Shimizu I, Sato T, Yoshida Y, Hayashi Y, Suda M, 等。γ-氨基丁酸在棕色脂肪组织中的信号传导促进肥胖的全身代谢紊乱。Cell Rep. 2018;24:2827–.e5.
[文章](https://doi.org/10.1016%2Fj.celrep.2018.08.024) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC1cXhslSnur3E) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=30208310) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Gamma-aminobutyric%20acid%20signaling%20in%20brown%20adipose%20tissue%20promotes%20systemic%20metabolic%20derangement%20in%20obesity&journal=Cell%20Rep.&doi=10.1016%2Fj.celrep.2018.08.024&volume=24&pages=2827-.e5&publication_year=2018&author=Ikegami%2CR&author=Shimizu%2CI&author=Sato%2CT&author=Yoshida%2CY&author=Hayashi%2CY&author=Suda%2CM)
- Constans C, Ahnine H, Santin M, Lehericy S, Tanter M, Pouget P, 等. 通过GABA输送穿越血脑屏障的非侵入性超声调制视觉诱发反应. J Control Release. 2020;318:223–31.
[文章](https://doi.org/10.1016%2Fj.jconrel.2019.12.006) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC1MXisVGis7fO) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=31816362) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Non-invasive%20ultrasonic%20modulation%20of%20visual%20evoked%20response%20by%20GABA%20delivery%20through%20the%20blood%20brain%20barrier&journal=J%20Control%20Release.&doi=10.1016%2Fj.jconrel.2019.12.006&volume=318&pages=223-31&publication_year=2020&author=Constans%2CC&author=Ahnine%2CH&author=Santin%2CM&author=Lehericy%2CS&author=Tanter%2CM&author=Pouget%2CP)
- Charopoulos I, Orme S, Giannoudis PV. 地舒单抗在骨质疏松症治疗中的作用和疗效:更新。专家意见药物安全。2011;10:205–17.
[文章](https://doi.org/10.1517%2F14740338.2010.516249) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BC3MXitlaru7o%3D) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=21208140) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=The%20role%20and%20efficacy%20of%20denosumab%20in%20the%20treatment%20of%20osteoporosis%3A%20an%20update&journal=Expert%20Opin%20Drug%20Saf&doi=10.1517%2F14740338.2010.516249&volume=10&pages=205-17&publication_year=2011&author=Charopoulos%2CI&author=Orme%2CS&author=Giannoudis%2CPV)
- Deeb SS, Fajas L, Nemoto M, Pihlajamäki J, Mykkänen L, Kuusisto J, 等. PPARγ2 中的 Pro12Ala 替代与受体活性降低、体重指数降低和胰岛素敏感性改善相关. Nat Genet. 1998;20:284–7.
[文章](https://doi.org/10.1038%2F3099) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DyaK1cXntFOhsL8%3D) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=9806549) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=A%20Pro12Ala%20substitution%20in%20PPAR%CE%B32%20associated%20with%20decreased%20receptor%20activity%2C%20lower%20body%20mass%20index%20and%20improved%20insulin%20sensitivity&journal=Nat%20Genet&doi=10.1038%2F3099&volume=20&pages=284-7&publication_year=1998&author=Deeb%2CSS&author=Fajas%2CL&author=Nemoto%2CM&author=Pihlajam%C3%A4ki%2CJ&author=Mykk%C3%A4nen%2CL&author=Kuusisto%2CJ)
- Gong Z, Huang C, Sheng X, Zhang Y, Li Q, Wang M-W, et al. Tanshinone IIA 在通过过氧化物酶体增殖物激活受体 γ 拮抗作用治疗肥胖中的作用。内分泌学。2009;150:104–13. 和 YQZ.
[文章](https://doi.org/10.1210%2Fen.2008-0322) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BD1MXmt1alug%3D%3D) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=18818299) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=The%20role%20of%20Tanshinone%20IIA%20in%20the%20treatment%20of%20obesity%20through%20peroxisome%20proliferator-activated%20receptor%20%CE%B3%20antagonism&journal=Endocrinology.&doi=10.1210%2Fen.2008-0322&volume=150&pages=104-13&publication_year=2009&author=Gong%2CZ&author=Huang%2CC&author=Sheng%2CX&author=Zhang%2CY&author=Li%2CQ&author=Wang%2CM-W)
- 陈 A, 科勒 AN. 转录因子抑制:经验教训与新兴靶点. 分子医学趋势. 2020;26:508–18.
[文章](https://doi.org/10.1016%2Fj.molmed.2020.01.004) [CAS](https://www.nature.com/articles/cas-redirect/1:CAS:528:DC%2BB3cXitlGrt74%3D) [PubMed](http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=32359481) [PubMed Central](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC7198608) [谷歌学术](http://scholar.google.com/scholar_lookup?&title=Transcription%20factor%20inhibition%3A%20lessons%20learned%20and%20emerging%20targets&journal=Trends%20Mol%20Med&doi=10.1016%2Fj.molmed.2020.01.004&volume=26&pages=508-18&publication_year=2020&author=Chen%2CA&author=Koehler%2CAN)
'下载参考文献'
作者感谢CRC核心设施提供的技术和方法支持。 “基因分型和生化中心(CGB)”的工作人员进行了基因分型。我们衷心感谢“功能探索中心(CEF)”和“rAAV平台”(法国Imagine研究所)的工作人员在体内实验中的技术支持。切片制备和图像采集由“组织学、细胞成像和流式细胞术中心(CHIC)”进行。卡通图像已从Biorender软件下载。GK得到了癌症对抗联盟(équipe labellisée);国家研究机构(ANR)——白色项目;AMMICa US23/CNRS UMS3655;癌症研究协会(ARC);“粉红丝带”协会;法兰西岛癌症中心;医学研究基金会(FRM);Elior的捐赠;Equipex Onco-Pheno-Screen;罕见疾病欧洲联合计划(EJPRD);古斯塔夫·鲁西奥Odyssea,欧盟地平线2020项目Oncobiome和Crimson;家乐福基金会;国家癌症研究所(INCa);Inserm(HTE);法国大学研究所;LabEx免疫肿瘤学(ANR-18-IDEX-0001);Leducq基金会;来自马克基金会的癌症研究ASPIRE奖;RHU都灵光;Seerave基金会;SIRIC分层肿瘤细胞DNA修复和肿瘤免疫消除(SOCRATE);以及SIRIC癌症研究和个性化医学(CARPEM)的支持。本研究贡献于IdEx巴黎大学ANR-18-IDEX-0001。GA得到了医学研究基金会(FRM)的支持。HC得到了中国国家留学基金委的支持。JMB-SP由“拉蒙·卡哈尔计划”(RYC-2018-025099-I)资助。
作者注释
- 这些作者共同监督了这项工作:José Manuel Bravo-San Pedro, Isabelle Martins 和 Guido Kroemer.
- 科尔德利研究中心,法国巴黎大学,索邦大学,法国国家健康与医学研究院 U1138,法国大学研究所,巴黎,法国
Gerasimos Anagnostopoulos, Omar Motiño, Sijing Li, Vincent Carbonnier, Hui Chen, Sylvère Durand, Mélanie Bourgin, Fanny Aprahamian, Nitharsshini Nirmalathasan, Konstantina Kotta, Léa Montégut, Flavia Lambertucci, Maria Chiara Maiuri, José Manuel Bravo-San Pedro, Isabelle Martins & Guido Kroemer
- 代谢组学与细胞生物学平台,古斯塔夫·鲁西研究所,维尔久夫,法国
Gerasimos Anagnostopoulos, Omar Motiño, Sijing Li, Hui Chen, Sylvère Durand, Mélanie Bourgin, Fanny Aprahamian, Nitharsshini Nirmalathasan, Léa Montégut, Flavia Lambertucci, Maria Chiara Maiuri, José Manuel Bravo-San Pedro, Isabelle Martins & Guido Kroemer
- 巴黎萨克雷大学医学院,克雷蒙比凯特,法国
Gerasimos Anagnostopoulos, 李思静, 陈辉 & Léa Montégut
- 实验与健康科学系,庞培法布拉大学(UPF),西班牙巴塞罗那;神经退行性疾病网络生物医学研究中心(CIBERNED),西班牙马德里
瓦伦蒂娜·西卡
- 增殖、压力与肝脏生理病理实验室,科尔德利研究中心,75006,巴黎,法国
罗曼·多纳 & 香塔尔·德斯杜
- 科尔德利研究中心,法国国家卫生与医学研究院,索邦大学,巴黎大学,F-75006,巴黎,法国
罗曼·多纳 & 香塔尔·德斯杜
- INSERM U1163,想象研究所,巴黎,法国
马塞洛·西蒙·索拉
- INSERM U830, Équipe Labellisée LNCC, 儿童肿瘤的多样性与可塑性实验室, PSL 研究大学, SIREDO 肿瘤中心, 纪念居里研究中心, 75005, 巴黎, 法国
Didier Surdez & Grossetête Sandrine
- 骨肉瘤研究实验室,巴尔格里斯特大学医院,苏黎世大学,苏黎世,瑞士
迪迪埃·苏尔德兹
- INSERM U830, Équipe Labellisée LNCC, 儿童肿瘤的多样性与可塑性实验室, PSL研究大学, SIREDO肿瘤中心, 纪尧姆·居里研究中心, 75005, 巴黎, 法国
奥利维耶·德拉特尔
- 体细胞遗传学单元,肿瘤遗传学服务,居里研究所,医院中心,75005,巴黎,法国
奥利维耶·德拉特尔
- 马德里大学医学院,生理学系,马德里,西班牙
何塞·曼努埃尔·布拉沃-圣佩德罗
- 生物学中心,乔治·蓬皮杜欧洲医院,AP-HP,巴黎,法国
圭多·克罗默
GK、IM 和 GA 设计了研究。GK 和 GA 撰写了论文。GA、OM、IM、VS、SL、HC、KK、LM、FL 和 RD 进行了哺乳动物细胞生物学和小鼠研究。SL、VC、IM 和 SL 进行了生物信息学分析。SD、FA、MB 和 NN 进行了代谢组学分析。MSS 生产了 AAV8 载体。IM、MCM、JMB-SP、CD、DS 和 OD 提供了智力支持并编辑了论文。所有作者审阅了结果,编辑并批准了手稿的最终版本。
GK与第一三共、Eleor、Kaleido、Lytix Pharma、PharmaMar、Samsara、赛诺菲、Sotio、Tollys、Vascage和Vasculox/Tioma保持着研究合同。GK是布里斯托尔-迈尔斯-斯奎布基金会法国的董事会成员,也是EverImmune、Samsara Therapeutics、Therafast Bio的科学共同创始人。GK是涵盖老化、癌症、囊性纤维化和代谢疾病的治疗靶向专利的发明人。GK、JMB-SP和OM是涵盖抗-ACBP/DBI抗体治疗用途的专利的发明人。GK是Osasuna Therapeutics的创始人,该公司专注于ACBP/DBI的靶向治疗。
- 分子癌症 (2024)
- 自然代谢 (2024)
- 细胞死亡与疾病 (2024)